东南亚人类古基因组学:从全新世到殖民时期的远距离扩散与社区结
古代基因组学(又称考古遗传学)领域自20世纪80年代初创以来取得了长足发展。在技术和方法创新的推动下,现在可以研究人类历史、病原体演化、环境DNA以及许多其他研究问题。 由于化学和生物因素的影响,古DNA通常是片段化的、化学修饰的,且数量稀少,因此需要特殊的实验室提取协议。在热带地区,提取工作更具挑战性,这也是为什么迄今为止从东南亚(一个具有丰富历史和高度文化与遗传多样性的地区)发表的古代基因组相对较少。作为本论文的一部分,对来自东南亚的一百多个化石骨骼样本进行了采样,这些样本的年代从中全新世到殖民时期不等。提取的古DNA在来自相同和不同遗址的个体间保存质量差异很大,其中岩骨通常比采样的牙齿

摘要
古代基因组学(又称考古遗传学)领域自20世纪80年代初创以来取得了长足发展。在技术和方法创新的推动下,现在可以研究人类历史、病原体演化、环境DNA以及许多其他研究问题。
由于化学和生物因素的影响,古DNA通常是片段化的、化学修饰的,且数量稀少,因此需要特殊的实验室提取协议。在热带地区,提取工作更具挑战性,这也是为什么迄今为止从东南亚(一个具有丰富历史和高度文化与遗传多样性的地区)发表的古代基因组相对较少。作为本论文的一部分,对来自东南亚的一百多个化石骨骼样本进行了采样,这些样本的年代从中全新世到殖民时期不等。提取的古DNA在来自相同和不同遗址的个体间保存质量差异很大,其中岩骨通常比采样的牙齿含有更多DNA。通过基因富集方法,生成了68个古代基因组,其中包括来自瓦拉西亚(Wallacea)的第一个也是最古老的基因组。
尽管现代人类最初抵达东亚的时间点仍有争议,但瓦拉西亚(位于大陆和巴布亚新几内亚之间的深海岛屿)的首次横越和定居已可靠地确定为至少在5万年前。在手稿A中描述的对一个7,200年前来自苏拉威西的个体的基因分析揭示了瓦拉西亚和大陆在中全新世期间或之前的基因接触。这个个体最好被模型化为与大洋洲和东亚人群相关的祖先谱系的混合,同时还携带约2.2%的丹尼索瓦人基因组,这加剧了关于华莱士线以外丹尼索瓦人基因渗入的争论。
随着水稻和粟作农业在11,000年前在现今中国的长江和黄河流域的发展,狩猎采集文化在东南亚被取代。沿河流向大陆东南亚以及乘船向岛屿东南亚的进一步技术转移带来了深刻的文化和人口变化。手稿B描述了对来自瓦拉西亚不同地区、年代在2,600至5,250年之间的个体的基因分析结果,证明了南部和西部瓦拉西亚地区的南亚语系和南岛语系相关群体之间的基因交流。
与新石器化过程相反,冶金术的起源和传播路线仍存在争议。其中,矿山、冶炼炉和工艺品的发现表明,铜在约3,000年前开始被加工,之后不久便出现了铁器。在泰国西北部,带有在头部和脚部雕刻的木质棺材标记了四十多个年代在2,300至1,000年之间的发掘地点。在手稿C中展示了相关人群含有狩猎采集者、长江和黄河相关的基因组成,这表明不同新石器时代谱系可能有不同的扩散路线和交流。对来自多个发掘地点的33个基因组的分析还显示,同一地点的个体之间存在密切的亲缘关系,而不同地点的个体之间存在较远的关系。
日益增长的贸易关系以及香料和陶瓷的可获得性从公元1世纪开始首先导致了沿海城市的国家形成,随后引起了欧洲的兴趣和征服。从16世纪开始,西班牙首先殖民了菲律宾的部分地区,然后是台湾北部。在台湾北部和平岛的考古发掘中,发现并研究了一座殖民时期教堂的地基和毗邻的墓地。手稿D描述了一个基因多样性的社区,其中欧洲血统的个体被埋葬在教堂围墙内,而东亚和东南亚血统的个体则被埋葬在教堂毗邻区域。这表明社会等级制度在墓葬排列中得到体现,即使在1642年被荷兰人征服之后仍然如此。
所提交的手稿证明,通过创新方法,可以从东南亚许多地区获取和分析古DNA,并展示了研究远距离基因交流和当地社区结构的潜力。为了不进一步加剧西方与当地研究人员之间的技术和财政不平衡,平等合作和当地社区参与是开发互惠互利的考古遗传学项目的重要前提,这些项目可以进一步探索东南亚的历史。
总结
自20世纪80年代初以来,古基因组材料(又称考古遗传学)研究领域发展迅速。在技术和方法创新的推动下,考古遗传学研究现在可以探索人类历史、病原体演化、环境DNA等众多领域。
由于化学和生物变异,古DNA通常是片段化的、化学修饰的,且数量极少,这需要特殊的实验室提取协议。在热带地区,这些挑战更加严峻,这也是为什么迄今为止从东南亚(一个人类历史丰富、现代文化和基因多样性高的地区)发表的古代基因组相对较少。作为本论文的一部分,我们对来自东南亚大陆和岛屿的一百多个考古个体进行了采样,这些样本的年代从中全新世到殖民时期不等。我们观察到古DNA在不同遗址内部和之间的保存情况差异很大,其中颞骨的岩骨部分通常比牙齿表现更好。利用溶液捕获技术,我们能够生成并验证来自68个古代人类的全基因组数据,其中包括来自瓦拉西亚的第一个也是最古老的测序个体。
虽然现代人类最初出现在东亚的时间点仍有争议,但瓦拉西亚(位于大陆和巴布亚新几内亚之间的海洋岛屿)的首次横越和定居已可靠地确定为至少在5万年前。手稿A中描述的来自苏拉威西岛Leang Panninge的一个7,200年前个体的数据揭示了在中全新世期间或之前与大陆的接触。这个个体最好被模型化为一个超过37,000年前与近大洋洲人相关的谱系和深层东亚相关祖先的混合,同时还携带约2.2%的丹尼索瓦人相关基因组,这加剧了关于华莱士线以外丹尼索瓦人基因渗入的争论。
该地区长期存在的狩猎采集者相关技术复合体被约11,000年前分别在长江和黄河流域发展起来的水稻和粟作农业所打断。这些技术沿河流向东南亚大陆传播,并通过船只向岛屿东南亚传播,与当地狩猎采集者的互动产生了深远的文化和人口影响。手稿B呈现了来自瓦拉西亚不同地区、年代在2,600至250年前之间的基因组数据,证明了南亚语系和南岛语系相关扩散的互动延伸到了西部和南部瓦拉西亚。
相比之下,冶金术在东南亚的起源和传播仍然存在争议。矿山、冶炼炉和工艺品表明,至少在3,000年前,青铜和随后不久的铁器已遍布东南亚大陆。在泰国西北部,具有独特头部和脚部雕刻的木棺标记了40多个年代在2,300至1,000年前之间的遗址。在手稿C中,我们展示了相关个体携带狩猎采集者、长江和黄河相关的祖先成分,这可能表明两种新石器时代相关祖先有不同的扩散路线和互动。通过对来自多个遗址的33个基因组的分析,我们还能够识别出遗址内部的密切基因关系和遗址之间的较远关系。
日益增长的贸易联系以及香料和陶瓷的可获得性首先导致了从公元1世纪开始的沿海政权的国家形成,随后引起了欧洲的兴趣和征服。从16世纪开始,西班牙殖民了菲律宾的部分地区,之后是台湾北部。在台湾北部和平岛的考古发掘中,发现了殖民时期教堂的地基和毗邻的墓地。手稿D描述了一个基因多样性的社区,其中欧洲相关血统的个体被埋葬在教堂内,而东亚和东南亚相关个体则被埋葬在教堂旁边。这表明社会等级制度在墓葬空间排列中得到体现,可能在1642年荷兰征服后仍然如此。
手稿D:和平岛殖民时期墓地的人口多样性
在K. Nägele和C. Posth的指导下,我进行了质量控制和人群遗传分析,绘制了所有图表,并撰写了手稿和补充材料,K. Nägele、M. Cruz Berrocal、F. Valentin和J. Krause提供了意见和建议。
目录
缩略语 4
德语摘要 5
英文摘要 7
出版物和手稿列表 9
已接受的手稿 9
已提交的手稿 9
准备提交的手稿 9
个人贡献 10
1. 考古遗传学和东南亚历史简介 11
1.1 考古遗传学的发展和当前方法 11
1.2 东南亚的古人类和现代人类 16
2. 研究目的和目标 32
3. 手稿 33
3.1 手稿A:来自瓦拉西亚的中全新世狩猎采集者基因组 33
3.2 手稿B:过去三千年的古代基因组支持多次人类向瓦拉西亚的扩散 54
3.3 手稿C:泰国西北部铁器时代木棺文化的基因组肖像和亲缘关系模式 71
3.4 手稿D:通过考古学、考古墓葬学和考古遗传学研究台湾早期殖民地点的占据历史、社会等级和生活史 89
4. 东南亚考古遗传学的潜力与挑战 106
4.1 克服具挑战性的DNA保存条件 106
4.2 追踪东南亚的定居过程和狩猎采集者多样性 109
4.3 阐明丹尼索瓦人基因渗入事件 111
4.4 描述区域连通性和交流 112
4.5 理解考古遗址层面的社会结构 117
4.6 发展公平的东南亚考古遗传学研究 121
5. 结论与展望 127
致谢 129
参考文献 130
手稿A-D的补充信息 160
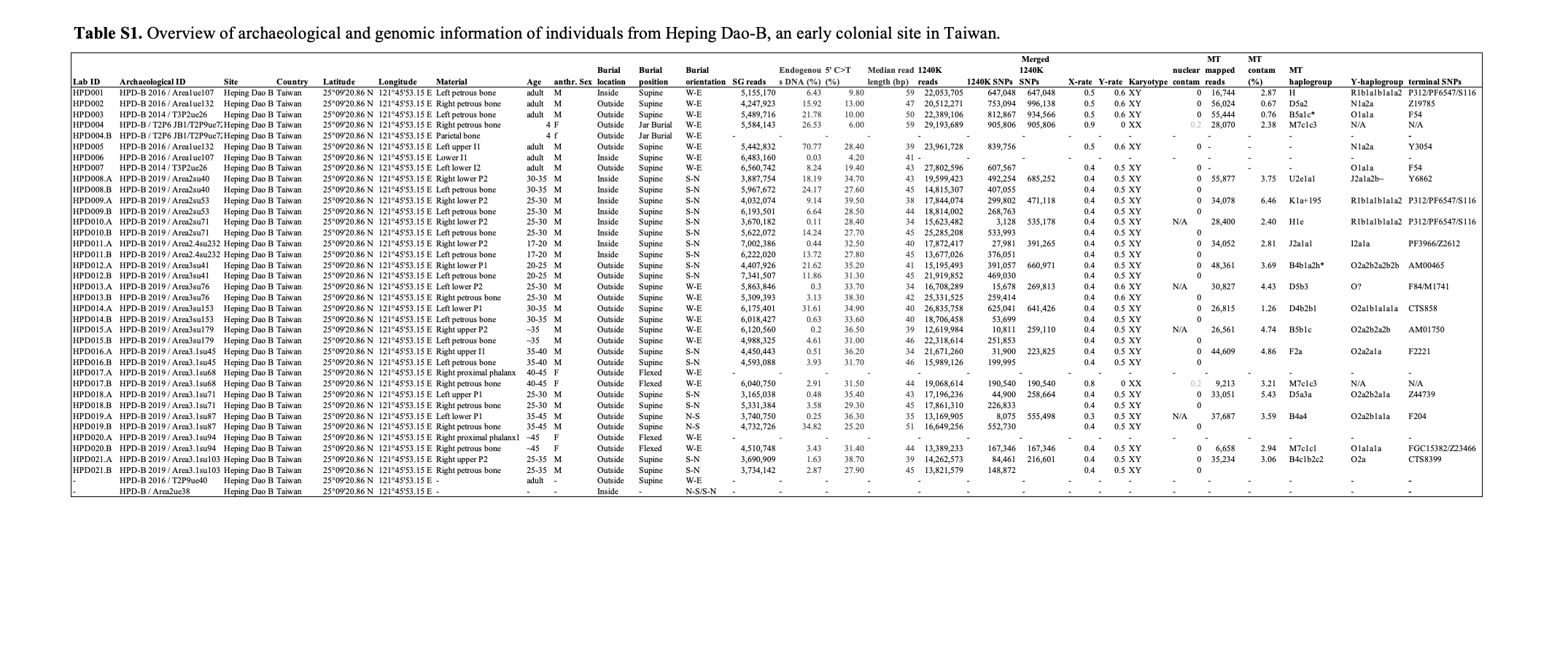
HPD002 HPD-B 2016 / Area1ue132 Heping Dao B Taiwan 25°09'20.86 N 121°45'53.15 E Right petrous bone adult M Outside Supine W-E 4,247,923 15.92 13.00 47 20,512,271 753,094 996,138 0.5 0.6 XY 0 56,024 0.67 D5a2 N1a2a Z19785
2011年至2019年期间,由M.C.B.、C.-h.T.和E.H.(自2019年起)领导的西班牙和台湾联合国际团队在和平岛B区共发掘了十九座墓葬(图1b,2c)。其中四座墓葬(Area1ue107-HPD001/6、Area1ue132-HPD002/5、T2P2ue26-HPD003/7、T2P9ue72-HPD004)之前已有详细描述11,12。在教堂基础内HPD001北侧,发掘了四座仰卧位成人墓葬(HPD008-11)和一个坑(Area2ue38)。与HPD001不同,这些墓葬呈南北向。在教堂外HPD002/5和HPD003/7南侧,我们发现了六座呈西东向的仰卧位成人墓葬。在这些墓葬南侧,有四座呈南北向的仰卧位成人墓葬和一座呈北南向的仰卧位成人墓葬。在这些墓葬的左右两侧,我们发现了两座呈西东向的侧身屈肢成人墓葬(右侧卧)和一座罐葬儿童墓(HPD004)。自2019年以来发掘的墓葬分析工作仍在进行中。
我们从和平岛18个个体的颞骨岩部和牙齿中提取了古DNA,这项工作在德国耶拿马克斯·普朗克人类历史科学研究所(现为马克斯·普朗克地球人类学研究所)进行。HPD001-7样本由F.V.采集,而S.C.和考古团队则在2019年原位采集了个体HPD008-21的岩骨和牙齿。在文库制备和鸟枪法筛选后,我们通过片段中值长度短和读段5'端胞嘧啶到胸腺嘧啶转换率升高来确认提取的DNA片段的真实性(图S1)。随后,我们使用溶液杂交捕获技术富集约1,240,000个单核苷酸多态性(SNPs)。
对整个人类基因组(1240K捕获17,18)进行了测序,每个个体的中位数为503,148个SNPs(SI表1)。我们确定了16个个体为XY染色体型,2个个体为XX染色体型,并获得了XY个体的Y-单倍型为O2a2b2a、O1a1a和R1b1a1b1a,而通过线粒体捕获19获得的线粒体单倍型则更加多样化(B、D、F、H、J、K、M、U)(SI表1,补充材料)。我们能够确认采样的牙齿和岩骨属于基因一致的个体,但未发现受测个体之间存在其他遗传关系(补充材料)。三个个体显示出较高的纯合片段,表明存在小群体规模(HPD004)和父母之间的近亲遗传关系(HPD001,HPD013)(图S2)。
为了开始人群遗传分析,我们对欧亚大陆的现代个体20-24计算了主成分分析(PCA)(图S3)。和平岛的五个个体与东南亚群体聚类,八个与东亚群体聚类,而另外五个个体则与西欧亚群体聚类。两个分别仅包含东亚和西欧亚群体的区域PCA显示了古代个体之间的进一步分离(图2a、b,图S4)。我们还可以通过形式为f3(Mbuti, 现代群体, 测试)的遗传漂变计算和ADMIXTURE比例来确认这些视觉上的关联(图S5,S7)。
在东亚地区,HPD004、HPD017和HPD020与台湾的现代原住民和古代个体聚集在一起,在qpWave20分析中与台湾1-8世纪的汉本遗址24的个体在基因上一致(SI表2)。与台湾多个原住民群体的现代基因组比较显示,这些古代个体与鲁凯族、泰雅族和达悟族25最为相似,在ADMIXTURE分析中没有显示额外祖源系数的证据(图S7)。个体HPD012和HPD015在PCA中与东南亚个体聚集。虽然HPD015与菲律宾的现代伊戈罗特人26一致,但HPD012在qpWave和ADMIXTURE中显示了不同的东南亚相关祖源(图S7,SI表2)。HPD002-3、HPD013-14、HPD016、HPD018-19和HPD021与现代汉族人24聚集,在qpWave20分析中与中国沿海福建省300年前的川云个体27在基因上一致(SI表2)。
在西欧亚PCA上,HPD008和HPD010与现代西班牙人聚集,靠近西班牙北部赫罗纳8-12世纪的个体28(图2b),在qpWave20分析中与后者在基因上一致(SI表4)。HPD011聚集并与现代
与当代西班牙北部和巴斯克人群聚集在一起(图S5)。HPD009聚集在10-16世纪格拉纳达(西班牙南部)的一个群体28中,类似地可以被建模为8-12世纪赫罗纳、北非和中东个体28的基因混合(图S9,SI表5)。HPD001在PCA上没有与先前发表的同时期伊比利亚个体聚集,而是与现代法国人、德国北部10-12世纪的一个个体以及荷兰7-11世纪的个体29接近。使用qpWave20,我们确定这个个体与荷兰7-11世纪的个体最为相似,这些个体先前已被确认具有荷兰和西班牙基因成分29,我们可以将其祖源建模为约80%的7-11世纪荷兰和约20%的8-12世纪赫罗纳(图S8,SI表3)。
讨论
在此,我们展示了33个与泰国西北部铁器时代独木棺文化相关个体的全基因组数据。低水平的纯合片段和高线粒体单倍型多样性表明高地生活着一个大型社区,这一点得到了该地区发现的大量独木棺遗址的支持18,21。
从人群遗传学角度看,与独木棺相关的个体在基因上表现出同质性,在主成分分析位置和混合成分方面变异很小(图2a,补充材料)。这种模式表明与基因上不同的社区有较早的基因接触,或者在最近混合的社区内有大量的基因交流。
泰国西北部的独木棺相关社区保留了来自和平文化相关群体的祖源,而中国南方8千年前的群体携带龙林相关祖源,广西和越南青铜时代的年轻群体则不携带任何一种狩猎采集者相关成分(表S4)。这表明迁入群体与当地狩猎采集者有多次互动,同时也有很少与当地混合的迁移活动,导致东南亚形成了多元化的(后)新石器时代基因景观。这一点进一步得到了一些独木棺相关洞穴的占据历史可追溯到晚更新世和早全新世的证实3,5,并强调了理解这些互动和转变何时以及如何发生的重要性。然而,该地区较差的DNA保存条件使得从更早时期提取DNA特别具有挑战性。
在区域层面,我们在泰国古代个体中至少识别出两种不同的祖源谱系。来自高地的独木棺相关个体显示出一个与黄河流域群体相关的较大附加基因成分,这一成分在泰国东北部班昌青铜时代(3.1千年前)和铁器时代(2.5千年前)个体中不存在10。这些基因结果表明铁器时代期间这些群体有不同的影响范围,这与泰国西北部、越南中部高地和中国南方的木棺埋葬相似性21相吻合,而与泰国东北部居住结构下的埋葬形式不同40。
此外,古蛋白质组学研究表明,来自Long Long Rak的独木棺相关个体的饮食严重基于C3植物,可能包括稻米和淡水鱼等食物,而泰国东北部新石器时代和铁器时代群体的饮食更加多样化,但C3食物来源较少41。尽管对线粒体DNA数据的分析得出结论认为独木棺文化是通过文化传播从中国云南省传入的17,但新生成的常染色体数据显示在独木棺引入泰国西北部之前或同时有外部基因影响。这种差异也可能反映了不同新石器时代群体的不同迁移路线,高地庞马帕是沿萨尔温河和山脉的西北路线的一部分。
研究的独木棺社区中携带黄河相关祖源的群体的基因流动在东亚和东南亚其他地区也有类似情况(图3a)。这一成分在中国福建省晚新石器时代之后的古代个体中被检测到9,在6.4到1.5千年前抵达中国广西省7,后来出现在马来西亚、缅甸、老挝和越南的新石器时代和青铜时代个体(4.2-2.1千年前)中7,8。最近发表的古代人类基因组数据还显示,上黄河相关祖源对一些早期和现代藏族高原群体贡献了>80%,表明在青稞农业引入高原之前就存在基因联系42。由于中国中原的现代群体在上黄河和东亚南部相关祖源之间形成基因渐变,前者被认为与汉藏语系语言从其上黄河沿岸的故乡在晚期磁山/早期仰韶时期(7.4千年前)向东南亚大陆的传播相关32,43。
泰国东北部的青铜时代和铁器时代班昌以及越南北部的新石器时代Man Bac在缺乏这一基因成分方面很突出,可能指向一种时间模式,即这一成分只是后来才到达泰国和越南,或者文化边界限制了进一步传播。结合新型混合建模和测年技术的额外古代基因组将有助于确定这一东亚北部成分的引入日期,并理解这一祖源在未来东亚和东南亚的分布和传播。
尽管对线粒体单倍群频谱的研究确定现代Khon Mueang和Thai Yuan与泰国西北部古代独木棺相关个体最为相似17,但在全基因组水平上,现代群体31对泰国西北部的古代个体显示出广泛的亲缘关系,但也携带额外的祖源成分(图3b,补充材料)。这表明1700年前后基因库发生了重大变化,包括历史时期来自老挝、缅甸和中国南方的迁移44,45。尽管如此,泰国西北部的群体仍保持与泰国其他地区的基因差异31,46,47。
从许多与同一考古复合体相关且在邻近区域发现的个体中生成基因组数据,也首次允许我们调查古代东南亚社区的社会结构。在独木棺相关遗址,我们发现了来自位于Mae Lana谷的Yappa Nhae 1和2的采样个体之间,以及在先前发表的来自Long Long Rak的基因组内部存在几个密切的基因关系。此外,位于Lang河谷的Lahu Pot和Tham Lod的个体,以及Yappa Nhae 2共享长IBD片段,表明不同河谷的社区之间存在紧密联系和交流。Yappa Nhae 2和Tham Lod的个体共享3至6度的亲缘关系,而Yappa Nhae 2和Lahu Pot则通过5至6度的亲缘关系更远地联系在一起。
该地区的现代群体通过单亲标记可追踪到母系和父系定居模式45,但对单亲标记的分析表明古代相关个体通过母系(例如,YPN014-YPN021)和父系(例如,YPN014-YPN027-YPN030)在Yappa Nhae 2内部有联系。Yappa Nhae和Long Long Rak内部的几个1级和2级关系,加上Yappa Nhae 2的六个个体和Tham Lod的一个个体彼此之间的关系比与遗址的其他个体更密切,可能表明基因亲属倾向于埋葬在同一洞穴中。这与基于棺木把手的社交网络分析相对应,该分析表明每个河谷的墓地遗址紧密相关,各自的社区跨河谷互动48,49。由于Long Long Rak的两个一级亲属个体也被埋葬在同一口棺材中8,棺木把手的多样性也可能反映了不同时期居住在高地庞马帕的家庭多样性。
将我们的考古基因学结果纳入考古发现,展示了对单一遗址和考古复合体进行深入研究的价值,以揭示更多关于过去社区的生活和信仰。对其他具有独木棺埋葬的遗址进行放射性碳测年和基因组分析将有助于进一步解析这些社区的社会结构,并研究跨区域联系和这种丧葬习俗的传播。
在东亚地区,HPD004、HPD017和HPD020与台湾现代原住民和古代个体聚类,并在qpWave20分析中与台湾1-8世纪汉本遗址的个体在基因上一致(SI表2)。与来自台湾多个原住民群体的现代基因组比较显示,这些古代个体与鲁凯族、泰雅族和达悟族25最为相似,且在ADMIXTURE分析中没有显示额外的祖源系数(图S7)。个体HPD012和HPD015在PCA分析中与东南亚个体聚类。虽然HPD015与菲律宾的伊戈罗特现代人群26一致,但HPD012在qpWave和ADMIXTURE分析中显示出不同的东南亚相关祖源(图S7,SI表2)。HPD002-3、HPD013-14、HPD016、HPD018-19和HPD021与现代汉族24聚类,并在qpWave20分析中与中国沿海福建省300年前的川云个体27在基因上一致(SI表2)。
图S3. 基于欧亚大陆现代个体32,33,35计算的主成分分析,和平岛的古代个体被投影并用灰色圆圈标记。为了单独关注这些聚类,我们计算了两个区域性PCA(图S4)。第一个PCA包括来自东亚和东南亚的个体(图S4A)。HPD004、HPD017和HPD020明显与台湾原住民群体聚类,而HPD012和HPD015则与菲律宾和马来西亚的个体聚类。HPD002-3、HPD013-14、HPD016、HPD018-19和HPD021与汉族人群重叠。
表格翻译:实验室ID 考古ID 遗址 国家 纬度 经度 材料 人类学年代 性别 埋葬位置 埋葬姿势 埋葬方向 SG读数 内源DNA(%) 5'C>T(%) 中位数读长(bp) 1240K读数 1240K SNPs 合并1240K SNPs X比率 Y比率 核型 核污染 线粒体映射读数 线粒体污染(%) 线粒体单倍群 Y染色体单倍群 末端SNPs
 陕公网安备61011302002223号
陕公网安备61011302002223号