东亚中新石器时代粟稻农业人群的基因组历史
[https://www.sciencedirect.com/science/article/pii/S2666979X25002320](https://www.sciencedirect.com/science/article/pii/S2666979X25002320) **The genomic history of East Asian Middle Neolithic millet- and rice-agricultural populations** **东亚中新石器时代粟稻农业人群的基因组历史** [1-s2.0-S2666979X25002320-mmc2.xlsx](%E

https://www.sciencedirect.com/science/article/pii/S2666979X25002320
The genomic history of East Asian Middle Neolithic millet- and rice-agricultural populations
东亚中新石器时代粟稻农业人群的基因组历史
1-s2.0-S2666979X25002320-mmc2.xlsx
研究亮点
- 中新石器时代黄河农民展示出明显的遗传亚结构
- 中新石器时代黄河与长江流域之间存在双向基因交流
- 最早的适应性EPAS1变体在黄河上游个体中被发现
- 原始南岛语族人群的遗传起源可追溯至更北方的长江流域
摘要
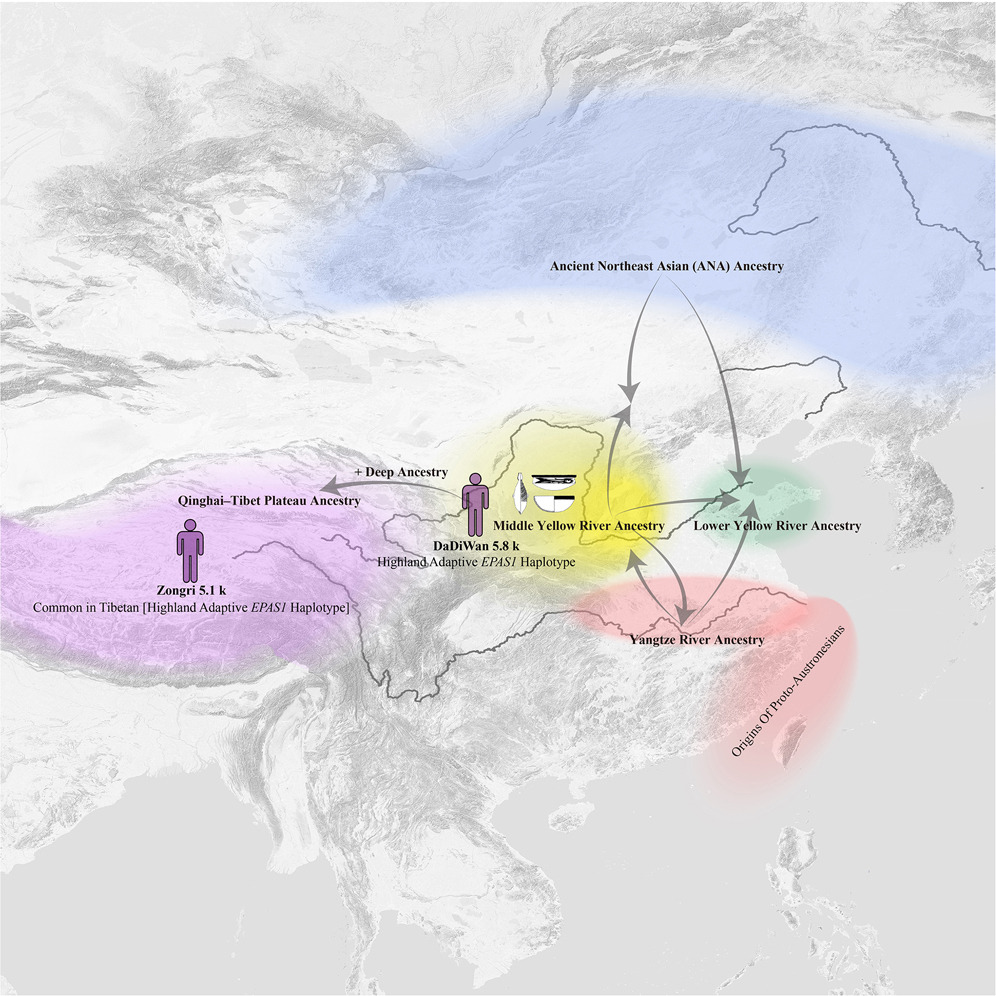
中国的黄河和长江流域是世界上最古老的独立农业中心之一,分别以粟和水稻的驯化而闻名,但它们的遗传历史尚未得到充分理解。在此,我们提供了来自这些地区的74个中新石器时代基因组样本的全基因组数据,显示出明显的遗传分化但存在双向基因流动,支持混合农业的人口扩散模型。黄河流域人群在中新石器时代粟农业扩张期间与周围群体的互动中呈现出明显的遗传亚结构。黄河上游人群与青藏高原人群在遗传上有联系,并拥有现代人类中最早的适应性EPAS1单倍型(约5,800年前)。同时,长江流域稻农与新石器时代至今的东南沿海中国和南岛语族人群显示出遗传亲缘关系,追溯原始南岛语族人群的起源至更北方的长江流域。这些发现为中新石器时代农业扩张对人类遗传历史的影响提供了新的见解
关键词
古DNA、中新石器时代、黄河与长江、原始南岛语族起源、适应性EPAS1单倍型
引言
农业的出现代表了从采集到耕作的重要转变,引发了生活方式、人口结构、健康状况、基因适应和文化等方面的深刻变革。1,2,3,4,5,6,7 这场农业革命在西亚、东亚和美洲等不同地区独立兴起,随后逐渐扩散到邻近区域。8,9,10,11,12,13 考古研究已经绘制了农业传播的时间线、地点和路线。然而,这种扩张背后的机制复杂且多方面,关于人口扩散和文化扩散的争论仍在继续。人口扩散理论认为人口迁移促进了农业的传播,而文化扩散理论则表明当地的狩猎采集者通过文化互动采纳了农业实践。14,15 古DNA分析提供了一种方法来研究农业传播是由人口替代、混合还是思想交流导致的。最近的研究揭示了不同地区农业扩张的多样模式。例如,在中部安纳托利亚,人类迁移在农业出现中扮演的角色微乎其微。16 相比之下,农业在整个欧洲的传播主要是由早期农民的扩张驱动的。17,18,19,20,21,22,23 值得注意的是,玉米农业从中美洲到美国西南部的传播是通过群体间扩散实现的,而非中美洲农民的长距离迁移。24 这些例子强调了农业实践与人口迁移之间的复杂关系。然而,对东亚这一重要的早期谷物作物驯化中心的这些动态进行全面探索的工作仍有待开展。
与西南亚和中美洲统一的农业系统不同,东亚呈现出双重农业结构:水稻(Oryza sativa)在中国南方占主导地位,而黍(Panicum miliaceum)和粟(Setaria italica)则在中国北方广泛种植。这两种作物约在10,000年前在长江流域和黄河流域被驯化,深刻塑造了中国的人口格局、健康状况和文化。黍和稻在早期中国文明的发展以及史前食物全球化过程中发挥了至关重要的作用。证据表明,黍的农业在7,000至5,000年前显著扩展到邻近地区。也有观点认为,汉藏语系的传播与这一农业扩张同时发生。随着农业成为主要食物来源,各种考古文化开始兴盛,包括红山文化(4,500-3,000 BCE)、仰韶文化(5,000-2,700 BCE)、大汶口文化(4,300-2,600 BCE)、大溪文化(4,300-3,300 BCE)和崧泽文化(4,000-3,300 BCE)。以黍作为中心的仰韶文化在中国北方尤为有影响力和主导地位。在仰韶文化的庙底沟阶段,以中原为核心的文化共同体形成,首次促进了中国大部分地区考古文化的整合与连接。这一时期标志着文化互动前所未有的加强。严文明提出了"中原为中心的双重花朵模式",认为中原是各文化区域之间的重要纽带。相比之下,张光直提出了"中国互动圈"的概念,强调区域文化具有独特特征,并以平等主义为特点。这些模型增进了我们对复杂文化交流的理解。然而,这些农业扩张和文化互动在多大程度上伴随着人口迁徙仍不确定。
大多数学者将研究重点放在中国的边疆地区和沿海地区,如新疆、西藏、东北、西南以及东部和东南沿海,揭示了显著的基因多样性和频繁的人口迁徙。相比之下,黄河和长江流域在新石器时期的人口历史仍未被充分探索。在黄河流域,研究主要集中在中游和下游地区。在黄河中游区域,来自河南省与仰韶文化相关的个体表现出基因同质性,并且对西辽河地区的人口产生了重大影响。仰韶文化人口的扩张导致西辽河地区农业活动增加。来自黄河下游山东的早期新石器时代个体显示出与新石器时代北方东亚人、西伯利亚人和藏族人的基因相似性。在中晚期新石器时代,与大汶口文化相关的个体受到了来自黄河中游农业人口东向扩张的影响,并表现出三重基因结构。这项研究支持了"中原中心双花图案模型"假说。然而,由于样本保存方面的挑战,长江流域的基因组研究很少。陶等人报告了来自长江上游的11个个体的基因组数据,时间从晚期新石器时代到青铜时代。这些个体与稻农相关的血统没有直接联系,而是与黄河农民表现出亲缘关系。这表明在晚期新石器时代,粟米农业通过人口扩散传入中国西南地区。
尽管东亚在农业起源的考古记录中具有重要意义,但迄今为止发表的全基因组古DNA数据仍然有限。这表明我们对农业和农民在新石器化过程中的作用的当前理解仅仅触及表面。为了解决这一研究空白,我们从20个考古遗址收集了74个样本,这些遗址年代为6,231-4,650年前,涵盖了整个黄河流域和长江中下游地区。这些样本代表了各种文化,包括仰韶文化(n = 62)、大汶口文化(n = 3)、马家窑文化(n = 2)、大溪文化(n = 4)、崧泽文化(n = 2)和良渚文化(n = 1)。通过将这些新样本与先前发表的数据相结合,我们旨在构建中新石器时期东亚人群的高分辨率遗传历史。我们新生成的数据使我们能够独立研究两个关键方面:(1)中新石器时期粟和稻农业人群的遗传结构,以及(2)这些人群与周围群体之间的互动关系,从而揭示农业传播的机制。
结果
中新石器时代黄河和长江流域农民的全基因组数据
为了探索东亚两个主要农业起源中心人群的遗传组成和发展轨迹,我们从74个古代个体中提取了DNA,这些个体分布在粟作物种植起源地黄河沿岸的各个地点,以及稻作发源地长江中下游地区。这项分析在1,240k位点组上产生了98,959至1,144,948个SNP位点(平均455,510个)(见表S1A)。在黄河流域,我们提供了来自上游(甘肃省,四个遗址)的21个古代基因组,来自中游(山西省,两个遗址;陕西省,七个遗址;河南省,三个遗址)的43个基因组,以及来自下游(江苏省,单一遗址,后与我们实验室构建的最大现有黄河下游古DNA数据集合并)的3个基因组。在长江流域,我们报告了来自中游(重庆,一个遗址)的4个古代基因组和来自下游(上海,两个遗址)的3个基因组。这些样本涵盖了整个黄河流域和长江中下游地区,碳十四测定年代范围为6,231至4,650校准年前,这一时期标志着东亚从狩猎采集向农业的重大转变。
我们最初关注的样本需满足以下标准:(1) 有明显的古代DNA损伤证据,(2) 通过至少两种独立方法确认的一致性别判定,以及(3) 现代人类污染水平低于3%,这是通过所有样本的线粒体方法和男性样本的X染色体方法确定的(见表S1A)。有十六个样本被识别为潜在污染,因为它们不符合这些标准。我们提取了这些样本中表现出古代DNA损伤特征的片段,并重新评估了它们的污染水平(表S1B)。我们只纳入了那些具有超过40,000个目标SNPs和至少一种可靠污染估计方法的样本,最终选择了12个样本。我们成功获得了70个个体的高质量遗传数据(表1和S1C)。然后,我们根据DNA损伤模式修剪了读数,确保所有读数的末端转换率低于3%,而那些特别与典型古代DNA损伤信号相关的读数低于6%。最后,我们使用三种不同方法(READ、TKGWV2和lcMLkin)评估了这70个样本的遗传亲缘关系,并识别出七对关系比二级亲属更近的个体。我们从每对亲缘关系中排除了覆盖率较低的个体,不再进行后续分析(表S1D)。
Table 1. Summary of 70 ancient samples passing filter thresholds, related to Table S1C
| Site | Location | Date range/mean (BP) | Sample size | SNP range (1,240k panel) |
|---|---|---|---|---|
| Fuquanshan | lower Yangtze River, Shanghai City | 5,454–5,419 | 2 | 133,547–191,266 |
| Maqiao | lower Yangtze River, Shanghai City | 5,301 | 1 | 1,089,643 |
| Daxi | middle Yangtze River, Chongqing City | 6,156–5,750 | 4 | 44,473–113,310 |
| Liulin | lower Yellow River, Jiangsu Province | 6,226–5,350 | 3 | 47,732–180,761 |
| Zhugang | middle Yellow River, Henan Province | 5,819–5,450 | 5 | 42,538–573,213 |
| Shuanghuaishu | middle Yellow River, Henan Province | 5,224–5,150 | 9 | 174,523–859,302 |
| Sunzhuang | middle Yellow River, Henan Province | 5,122–5,000 | 3 | 145,232–241,704 |
| Zhencheng | middle Yellow River, Shanxi Province | 5,150–4,910 | 2 | 169,838–248,524 |
| Yuancun | middle Yellow River, Shanxi Province | 4,650 | 1 | 145,857 |
| Nanzhaizi | middle Yellow River, Shaanxi Province | 6,230 | 1 | 256,244 |
| Wuzhong | middle Yellow River, Shaanxi Province | 6,150–6,045 | 4 | 264,929–760,839 |
| Dongying | middle Yellow River, Shaanxi Province | 5,593–4,860 | 2 | 518,583–656,334 |
| Huipoling | middle Yellow River, Shaanxi Province | 5,510 | 1 | 623,779 |
| Yangguanzhai | middle Yellow River, Shaanxi Province | 5,250–5,122 | 9 | 137,901–968,818 |
| Matengkong | middle Yellow River, Shaanxi Province | 5,170 | 1 | 328,388 |
| Wangyangpan | middle Yellow River, Shaanxi Province | 4,692 | 1 | 650,659 |
| Gedachuan | upper Yellow River, Gansu Province | 6,231–5,537 | 9 | 122,692–772,789 |
| Wangjiayinwa | upper Yellow River, Gansu Province | 5,823–5,662 | 8 | 222,933–878,094 |
| Dadiwan | upper Yellow River, Gansu Province | 5,819–5,808 | 2 | 806,819–1,144,948 |
| Shannashuzha | upper Yellow River, Gansu Province | 5,169–5,124 | 2 | 98,959–188,080 |
我们收集了66个高质量、无关联的古代基因组(见表S1和图1A),平均包含436,669个来自1,240k面板的SNPs。这个数据集填补了我们知识中的许多空白,代表了这两个东亚农业中心最大且最全面的新石器时代基因组收集。我们首先进行了主成分分析(PCA),以探索黄河和长江地区人群的整体基因组结构(图1B和S1A)。第一主成分明确区分了南方和北方人群。来自长江的核心个体,特别是来自中长江的J31902和来自下长江的基因同质群体(I52901、J72907和J81323;在qpWave中p > 0.05,以下称为"China_Lower_Yangtze_River_MN"),在PCA图的南方部分紧密聚集,与来自中国东南部的人群如台湾_良渚_晚旧石器时代、中国_福建_七贺_细石器时代和中国_东南亚_沿海_早期新石器时代一起(图1B)。相反,所有来自黄河的人群都位于PCA图的北方部分,特别是沿着藏缅语系路线,该路线从现代汉族人延伸到青藏高原的现代和古代居民(图1B)。值得注意的是,黄河人群在PCA图中的位置反映了他们的地理分布:上游黄河人群与高地群体紧密聚集,而下游黄河人群则更接近现代汉族人。中游黄河人群位于这两个群体之间,表明根据沿河各自地区存在明显的亚结构。这些发现得到了无监督ADMIXTURE分析的进一步支持。尽管与浅绿色相关的成分在所有黄河人群中都很普遍,但从山西省到上游地区的人群在ADMIXTURE图中显示出与藏缅语系或青藏高原(以下简称高地)人群相关的基因成分(蓝色表示)。相比之下,这些成分在河南省到下游地区的人群中几乎不存在(图S1B)。
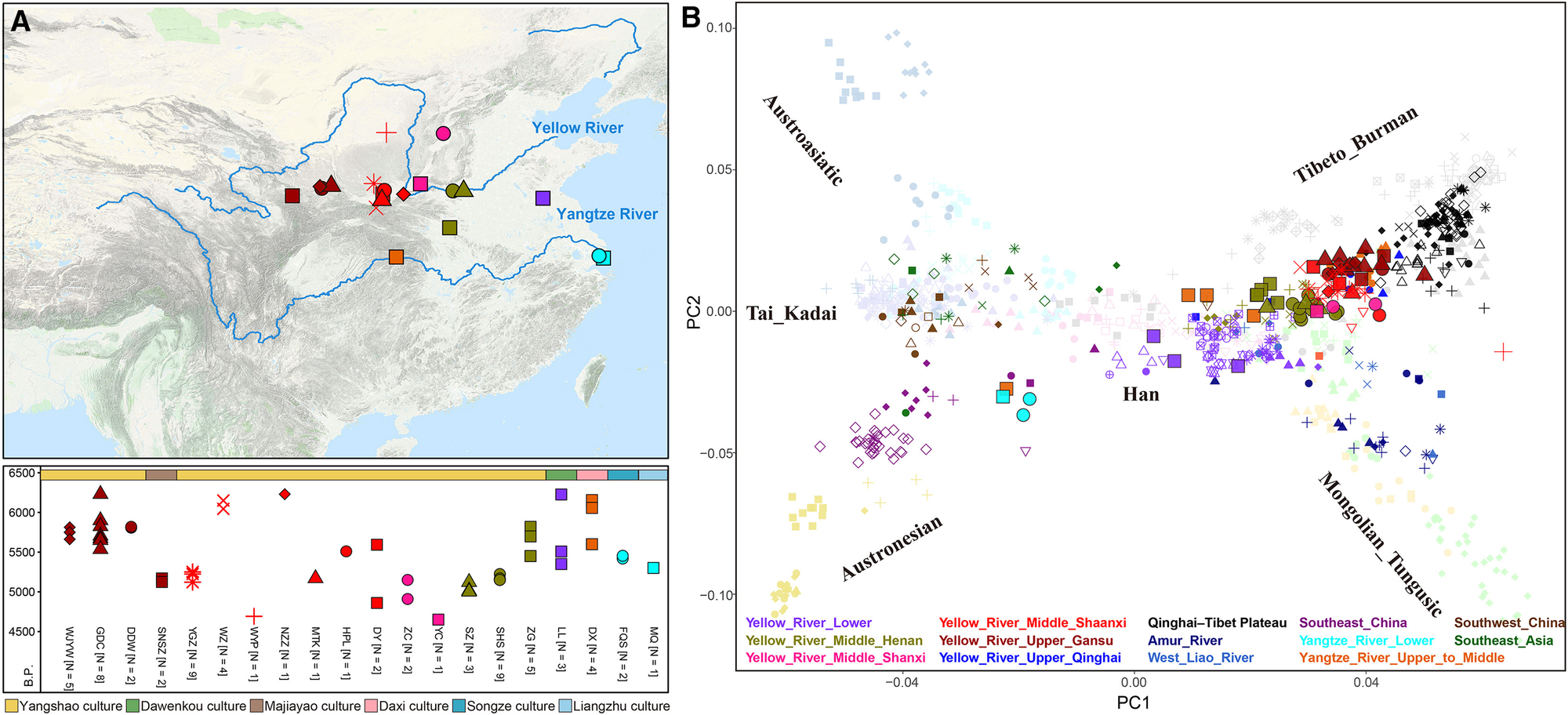
图1. 我们新采样群体的地理分布和人群结构
(A) 用于人群遗传分析的新采样群体的地理分布和日期。
(B) 现代东亚和东南亚个体的主成分分析(PCA)以及古代个体在PCA图上的投影。深色表示古代人群,浅色表示现代人群。较大的点表示我们新报告的个体。
黄河不同河段农耕者之间的遗传亚结构
先前的分析,包括主成分分析(PCA)和ADMIXTURE分析,已揭示出黄河特定区域人群之间存在明显的亚结构分化。然而,造成这种遗传亚结构的原因仍不清楚。为深入研究这一问题,我们利用基于旋转的qpWave分析将人群分为22个不同群体(见表S1E)。随后,我们进行了f4分析,格式为f4(Mbuti,参考人群;黄河人群,China_YR_MN),定量比较黄河各区域人群与China_YR_MN参考集的基因组特征(图S2A)。该分析纳入了13个具代表性的古代亚洲人群66作为参考,以识别促成所观察到的遗传亚结构形成的祖源人群和互动。
我们定性分析中发现的遗传亚结构进一步得到f4分析的证实,该分析表明与特定地理区域相关的人群与不同祖源人群表现出不同的遗传亲缘关系(图S2A)。关注河南省黄河中游地区,我们观察到该区域人群与China_YR_MN具有强烈的遗传相似性,其大部分(约90%)甚至全部(100%)祖源都可追溯至China_YR_MN(图2和S2B;表S2A)。这种区域性遗传特征既向东延伸至黄河下游西部(以Xixiahou_L和Ercun_ML为代表),也向西延伸到陕西东部和山西西南部(以Yangshao_YuanCun和Yangshao_NanZhaiZi为代表)。根据成对qpWave分析,这四个人群都与China_YR_MN在遗传上具有同质性(秩0:p > 0.05)。它们可以有效地以China_YR_MN作为唯一来源进行建模(图2和S2B;表S2A),表明该地区相对的遗传同质性和来自其他人群的影响最小。
相比之下,位于更东部和更西部的人群表现出不同的遗传模式。位于黄河下游东部的人群显示出来自当地狩猎采集者和南方人群的额外遗传贡献(图S2A)。这些人群最适合用二元、三元或四元混合模型来描述,反映了来自当地狩猎采集者、黄河中游农民(以China_YR_MN为代表)以及中国南方和古东北亚(ANA)人群的不同祖源比例。具体而言,它们的遗传构成包括23.8%–46.8%的China_NEastAsia_Coastal_EN相关祖源,24%–73.1%的China_YR_MN相关祖源,7.1%–57.3%的南方相关祖源,以及5.2%–16.5%的ANA相关祖源(图2和S2B;表S2A)。
相反,更西部的人群,从黄河中游到上游,相比其他群体展现出与高地人群的显著遗传亲缘关系(图S2)。这些人群被建模为二元或三元混合模型,具有来自黄河中游、高地和ANA人群的不同祖源贡献。它们的遗传构成包括21%至87.9%的China_YR_MN相关祖源,8.3%至55%的Zongri5.1k相关祖源,以及6.5%至91.7%的ANA相关祖源(图2和S2B;表S2A)。值得注意的是,黄河下游的人群含有更高比例的南方相关祖源(平均为32.3%),同时具有来自当地狩猎采集者的独特遗传贡献。相比之下,黄河中上游的人群独特地表现出Zongri5.1k相关祖源,这在黄河其他地区是不存在的。这种遗传差异可能直接解释了黄河农民之间观察到的亚结构。

图2. 新石器时代过渡期东亚的基因结构和混合
使用qpAdm模型估计新石器时代东亚人群的遗传构成。
虽然qpAdm的结果清晰直观,但我们也考虑了分析中可能存在的模型选择偏差。为了进一步验证我们的假设,我们检验了两种相关关系(图3A-3C和S3A)。第一种相关关系显示f4(Mbuti,China_NEastAsia_Coastal_EN/China_Lower_Yangtze_River_MN/China_SEastAsia_Coastal_LN;Yellow River populations,China_YR_MN)与经度之间存在显著的负相关(Pearson相关系数 = -0.777/-0.785/-0.710,p < 0.05),而与纬度没有显著相关(p = 0.073/0.643/0.998)(图3A、3B和S3A)。第二种相关关系显示f4(Mbuti,Zongri5.1k;Yellow River populations,China_YR_MN)与经度之间存在显著的正相关(Pearson相关系数 = 0.420,p < 0.05),而与纬度没有显著相关(p = 0.235)(图3C)。这些相关关系清楚地说明了上游黄河人群与高地人群之间以及下游黄河人群与南方人群和当地狩猎采集者之间的强烈基因亲缘关系,从而为我们的假设提供了定量验证。我们的研究结果表明,黄河流域不同地区的粟农之间存在明显的遗传亚结构。这种亚结构与地理分布相关,并受到与邻近人群互动的影响。
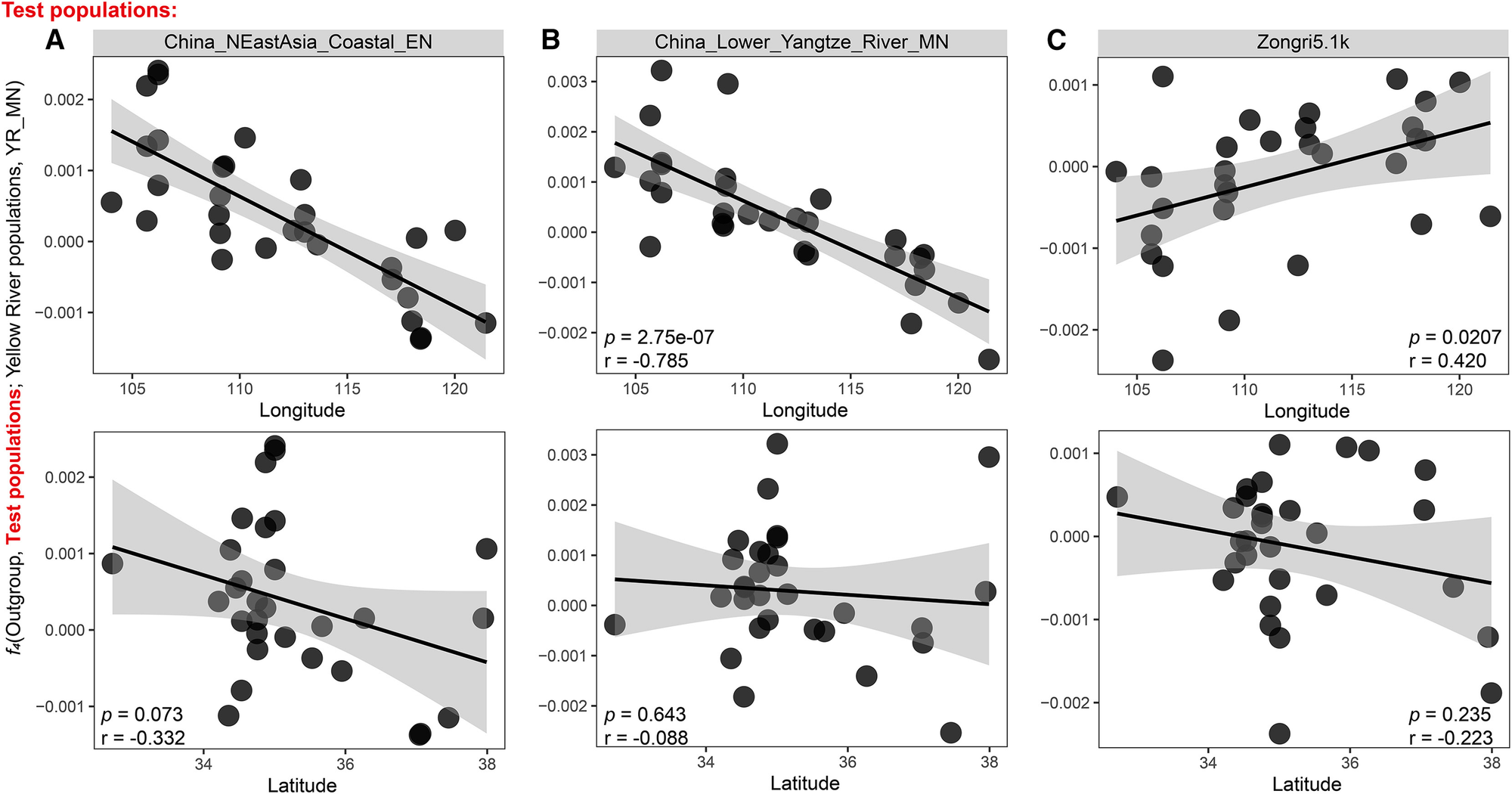
图3. 古黄河人群的f4统计值与纬度或经度相关性的估计
f4统计值以f4(Mbuti, 测试群体; 黄河人群, China_YR_MN)的形式呈现,其中测试群体代表(A) China_NEastAsia_Coastal_EN, (B) China_Lower_Yangtze_River_MN, 或(C) Zongri5.1k。每个点代表一个黄河人群,对应于图S2A中的行。p < 0.05被认为具有统计学显著性。
青藏高原祖先的主要来源追溯至黄河上游农民
现代人在青藏高原的永久定居被理论化为源自黄河地区的农业扩张,特别是涉及来自黄河中游至上游地区的农民。67,68 然而,这些地区古代基因组的缺乏限制了直接检验这一假设的能力。在本研究中,我们呈现了来自黄河上游(甘肃省,四个遗址)的17个中新石器时代基因组和来自中游(陕西和山西省,九个遗址)的22个中新石器时代基因组。这一地区此前被认为是高原人群的来源,而我们的数据代表了这些地区最早可获得的基因组。
我们的分析揭示,这些地区的人群在主成分分析(PCA)图中紧密聚集,与高原人群的亲缘关系更近(图1B)。ADMIXTURE分析图(图S1B)也识别出一个与高原人群相关的蓝色成分,这一成分在这些地区的人群中存在。这些发现进一步得到了定量f4分析和qpAdm分析的证实,表明只有这一地区的人群拥有Zongri5.1k相关祖源,这与其他黄河地区形成鲜明对比(图2和3C)。值得注意的是,来自甘肃省的Yangshao_GeDaChuan_2样本与高原人群表现出强烈的遗传联系,这一点从其在PCA图中的位置(图1B)以及f4(Mbuti, Zongri5.1k; Yangshao_GeDaChuan_2, 其他黄河中上游人群)的平均Z值为−3.19(图S4A)得到证实,并且估计其拥有超过50%的Zongri5.1k相关祖源(53.1%),这凸显了黄河上游与高原人群之间的遗传联系。此外,f4(Mbuti.DG, Zongri5.1k; 黄河人群, China_YR_MN)与经度之间观察到显著的正相关(皮尔逊相关系数 = 0.441,p < 0.05),而与纬度没有显著相关(p = 0.235)(图3C)。这提供了有力证据,表明高原祖源的主要来源可以追溯到黄河上游的农民,而非沿河的其他地区。
我们的分析显示,这些地区的人口在主成分分析(PCA)图中紧密聚集在一起,与高地人口的关系更为接近(图1B)。ADMIXTURE分析图(图S1B)也识别出了一个与高地人口相关的蓝色成分,这一成分在这些地区的人口中也有所体现。这些发现进一步得到了定量f4分析和qpAdm分析的证实,表明只有这一地区的人口携带了Zongri5.1k相关的祖源成分,这与其他黄河地区形成了鲜明对比(图2和3C)。值得注意的是,来自甘肃省的Yangshao_GeDaChuan_2展示了与高地人口的强烈基因联系,这一点从其在PCA图中的位置(图1B)以及f4 (Mbuti, Zongri5.1k; Yangshao_GeDaChuan_2, 其他中上游黄河人口)的平均Z值为−3.19(图S4A)得到了证实,并且估计其拥有超过50%的Zongri5.1k相关祖源成分(53.1%),这突显了上游黄河与高地人口之间的基因联系。此外,f4 (Mbuti.DG, Zongri5.1k; 黄河人口, China_YR_MN)与经度之间观察到显著的正相关(皮尔逊相关系数 = 0.441,p < 0.05),而与纬度没有发现显著相关(p = 0.235)(图3C)。这提供了有力证据,表明高原祖源的主要来源可以追溯到上游黄河地区的农民,而非黄河沿岸的其他地区。
有趣的是,同样位于甘肃省黄河上游的Yangshao_DaDiWan_1,是具有超过50%的Zongri5.1k相关祖先成分的个体之一(表S2A)。这个个体展示了与藏族相关的EPAS1(内皮PAS结构域蛋白1)单倍型的强烈信号(表S3A和S3B),这种单倍型是高原人群所独有的,有助于他们对高海拔环境的显著适应。这种单倍型被认为是由丹尼索瓦人引入的。68,69 虽然独特的藏族EPAS1单倍型已在可追溯至约5,100校准年前的高原个体中被识别出来,但在黄河沿岸的人群中却不存在。f4 (Mbuti, Yangshao_DaDiWan_1; Yangshao_GeDaChuan_1/Yangshao_GeDaChuan_4/Yangshao_WangJiaYinWa/China_Upper_YR_LN, highland populations)的平均Z值为−4.612(图S4B),而f4 (Mbuti, Zongri4.5k_o1; Yangshao_DaDiWan_1, Zongri4.5k/Zongri4.5k_o2/Zongri4.7k/Zongri5.1k)的平均Z值为5.62。这些结果与Yangshao_DaDiWan_1为藏族后裔的观点相矛盾,支持了这一结论:该个体实际上是黄河农民。Yangshao_DaDiWan_1的年代为5,819校准年前,代表了最古老的现代人类,也是首个已知携带高海拔适应性EPAS1等位基因的黄河农民。
黄河农民与古东北亚人群在高纬度地区的互动
古东北亚人群(ANAs)居住的区域跨越了远东西伯利亚、中国东北平原、蒙古高原和贝加尔湖地区。我们的研究结果表明,黄河流域的人群受到了古东北亚群体的显著影响(图2)。然而,目前尚不确定这种影响是集中在特定区域还是更广泛地分布在整个黄河流域,这需要进一步严格的研究。
为了解决这个问题,我们将与我们分析同时期的仰韶文化人群沿黄河分为三个地理组:上游(甘肃省)、中游(内蒙古、陕西、山西和河南省)和下游(山东和江苏省)。我们的研究结果显示,这些群体与ANA人群的遗传亲缘关系没有显著差异(p > 0.05,Wilcoxon检验)(图S5A)。值得注意的是,f4(Mbuti,Mongolia_East_N;黄河人群,China_YR_MN)的前三个最负Z值——出现在Yangshao_WangYangPan、China_Miaozigou_MN和Yangshao_ZhenCheng——都位于黄河中游高纬度地区(图S5B)。这表明ANA和黄河人群之间的互动在这个高纬度地区更加频繁。因此,我们从之前的中游组中提取这三个人群,形成一个新的类别,称为"高纬度组",并将其与黄河低纬度地区的人群进行比较。所有比较都显示出显著差异(p < 0.05,Wilcoxon检验)(图S5C)。鉴于Yangshao_WangYangPan和Yangshao_MaShanZhuGang在各自组中是异常值,我们将它们从分析中排除。为减轻高纬度组样本量有限的问题,我们纳入了一个后续时期的人群,即China_Shimao_LN。即使经过这种调整,显著差异仍然存在,进一步支持了我们的假设(p < 0.05,Wilcoxon检验)(图S5D)。
我们还进行了相关性测试,以正式评估与ANA人群的遗传亲缘关系与地理坐标(纬度和经度)之间的关系。我们的分析发现,f4(Mbuti,Mongolia_East_N;黄河人群,China_YR_MN)与纬度之间存在显著的负相关,皮尔逊相关系数为−0.613,p值小于0.05。然而,与经度没有发现显著相关(p = 0.761)(图S5E和S5F)。即使在排除先前确定的异常值后,这些结果仍然保持一致(图S5G),这直接证明了ANA人群对黄河流域的选择性影响,特别是在较高纬度地区。
此外,先前的研究表明,黄河农民的扩张可能促进了东北亚西辽河流域的农业发展。62,70 我们的分析表明,中新石器时代西辽河人群(China_WLR_MN)的基因组成可以被模拟为约50%的黄河人群(China_YR_MN + China_NEastAsia_Coastal_EN)和50%的ANA祖先成分(图2;表S2A)。相比之下,来自同一地区的China_HMMH_MN人群似乎完全源自ANA祖先成分(图2;表S2A)。这些发现表明,黄河农民可能对东北中国平原的ANA人群产生了重大影响,特别是通过农业实践的传播。总之,我们的结果强调了黄河农民与ANA人群之间的相互基因交流,这种交流可能在中国较高纬度地区更为频繁。
长江流域的遗传结构及其对南岛语族和黄河人群的影响作
为东亚水稻驯化的独立农业中心之一,长江流域的遗传历史由于缺乏长江农民的代表性古基因组而仍然难以解明。2023年,陶等人报告了来自上游长江流域的11个个体的基因组数据,这些个体的年代从新石器时代晚期到青铜时代,研究显示没有发现与水稻种植相关的祖先证据。为了解决这个问题,本研究提供了来自长江中游(大溪文化)的四个基因组和长江下游(崧泽和良渚文化)的三个基因组,它们可以追溯到中新石器时代。
长江流域核心人群(大溪_大溪_1、崧泽_福泉山和良渚_马桥)在主成分分析(PCA)图中与中国东南部新石器时代人群表现出遗传相似性(图1B和S1A)。它们表现出遗传同质性(秩0:p > 0.05,qpWave分析)并与中国东南部群体共享密切的祖先关系。这种关系进一步得到最大似然树(IQ-TREE)的支持,其中长江流域人群与东南沿海和岛屿人群聚集在一起,形成一个具有超过80%自展支持率的分支(图4A和S3F)。然而,与中国东南部群体相比,新石器时代长江流域人群与黄河流域人群共享更多等位基因,这一点在南方特异性PCA面板中表现为位于中国东南部和黄河流域聚类之间的中间位置(图S1C)以及f4(Mbuti,黄河流域人群;长江流域人群,中国东南部人群)显著为负的Z值(几乎所有Z < -3)(图S3G)。为了量化这些关系,我们应用了qpAdm,结果显示这些人群约60%的祖源来自中国东南部,约40%来自黄河流域(表S2B)。
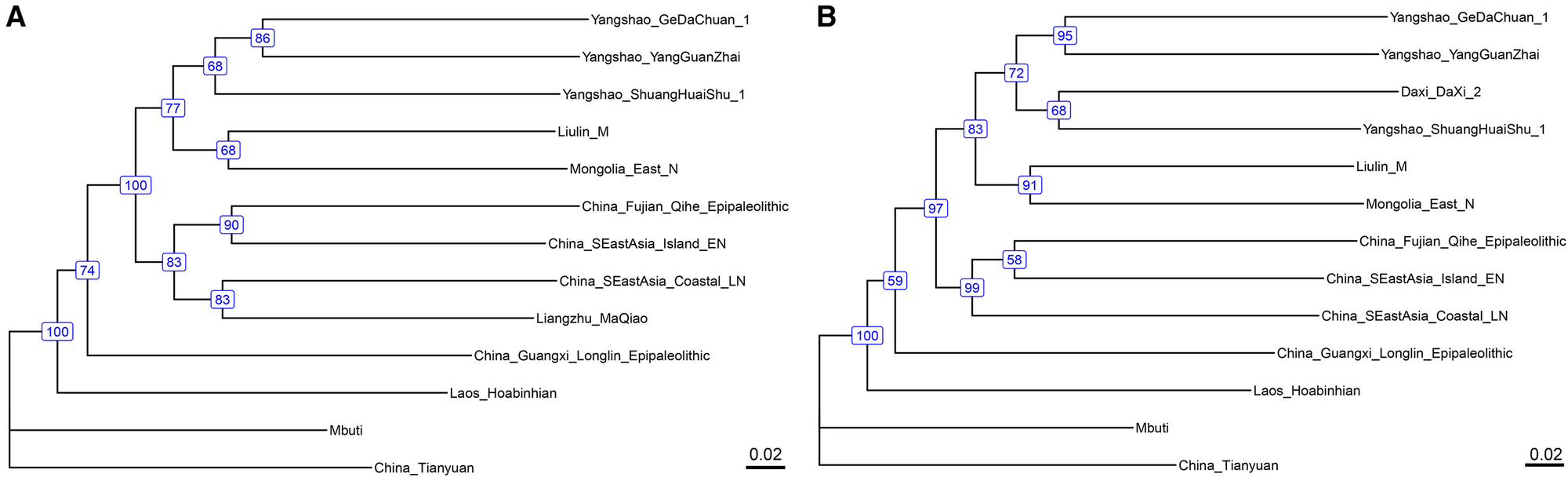
图4. 长江流域农民的系统发育关系
使用IQ-TREE估计的一致系统发育树拓扑结构,(A)显示了长江流域核心人群和(B)显示了大溪_大溪_2。
虽然长江流域的核心人群(大溪_大溪_1、崧泽_福泉山和良渚_马桥)表现出基因同质性并且主要祖源来自中国东南部,但中游长江(大溪_大溪_2)的三个个体显示出明显的基因差异。这些个体形成了一个与长江核心群体截然不同的集群,并在PCA图(图1B和S1C)和系统发育树(具有83%的自展支持率)(图4B)中表现出与北方人群更接近的基因亲缘关系,经qpAdm量化显示比核心群体多出11.5%的北方祖源成分和约5%的深层祖源成分(图2;表S2C)。在大溪_大溪_2中检测到少量(约5%)的深层祖源成分,但解释受到限制,原因是:(1)高损伤序列的选择性保留和(2)低覆盖度(平均约80,000个SNP位点)。如果这一信号是真实的,它可能来自附近的青藏高原或中国西南地区。更准确推断祖源成分将需要未来获得更高质量的基因数据。总体而言,这些结果表明长江中游人群可能经历了更复杂的人口互动。这些发现支持了粟米和水稻种植起源于不同祖先人群的假设。长江文明虽然与中国东南部群体有着强烈的基因亲缘关系,但已经受到了来自黄河地区人群扩张的影响。
此外,我们发现了这些人群之间相互影响的证据。下游黄河人群与南方人群的基因亲缘关系比其他地区更强,这从它们在主成分分析图(图1B)中的位置以及与南方人群的基因亲缘性和经度之间的显著正相关(图3B和S3A)得到证实。这种模式可能源于与长江流域人群的直接互动,而非来自东南或西南的新石器时代人群。相关性分析支持了这一结论,显示与其他同时期东南中国人群相比,China_Lower_Yangtze_River_MN表现出最负的基因相关系数(r = −0.785)(图3B和S3A–S3E)。qpAdm模型分析进一步证实,长江流域人群而非东南中国来源,是新石器时代山东群体唯一具有统计学支持的南方祖先贡献(表S2A;图2)。在河南省黄河中游地区,位于黄河和长江之间的Yangshao_MaShanZhuGang人群显示出14.7%的南方祖先成分,这也与长江流域起源有关(图2和S2B;表S2A)。这一发现表明,黄河中游地区的南方成分并非起源于晚期新石器时代,混合早在5,819年前(校正后年代)就已开始。这些发现表明,黄河中游和下游都受到了中新石器时代长江流域人群扩张的影响,而在黄河上游则没有这种贡献。我们的基因研究结果表明,这些地区粟米和水稻的共同生产伴随着直接且相互的人口流动和基因交流。
南岛语系人群分布遍及东亚,从台湾到西南太平洋,甚至延伸至东非的马达加斯加。有假设认为这些人群起源于中国大陆南部,但由于缺乏直接的古DNA比较,华南东南部与更北部长江流域的具体贡献仍不明确。我们的分析显示,现代台湾南岛语系人群与长江流域农民的基因亲缘关系比新石器时代晚期之前的东南沿海人群更为密切。这一点得到了f4统计量(Mbuti, Ami/Atayal; China_Lower_Yangtze_River_MN, China_Fujian_Qihe_Epipaleolithic/China_SEastAsia_Coastal_EN/China_SEastAsia_Island_EN)平均Z值为−5.30的支持。即使与新石器时代晚期的东南沿海人群相比,f4(Mbuti, Ami/Atayal; China_Lower_Yangtze_River_MN, China_SEastAsia_Coastal_LN)的Z值分别为−1.09和−0.89,进一步表明长江流域农民与现代台湾南岛语系人群之间存在密切的基因关系。基于这些发现,我们正式构建了qpAdm模型来确定现代南岛语系人群的基因构成。我们的结果表明,这两个人群最好用长江流域农民和台湾本地人群的组合来建模。具体而言,阿美族(Ami)的基因构成包括64.9%的长江流域相关祖源,27.5%的Taiwan_Hanben_IA相关祖源,以及7.6%的Taiwan_Liangdao_LatePaleolithic相关祖源。泰雅族(Atayal)的基因构成被模拟为55.9%的长江流域相关祖源和44.1%的Taiwan_Hanben_IA相关祖源。此外,Taiwan_Hanben_IA可以用三路模型建模,包含45.5%的长江流域相关祖源,48.1%的China_SEastAsia_Coastal_LN相关祖源,以及6.5%的Taiwan_Liangdao_LatePaleolithic相关祖源。这些结果表明,来自长江流域的基因流显著塑造了这些台湾南岛语系人群的基因库。总之,我们的发现支持这一假设:台湾原始南岛人的起源不仅与中国东南沿海有关,还与更北部地区,特别是长江流域有关。
讨论
文化协同与地理:塑造黄河农民的遗传亚结构
在本研究中,我们分析了来自中新石器时期的74个古基因组样本,这些样本跨越黄河和长江流域。我们的发现揭示了黄河农民中存在明显的遗传亚结构。值得注意的是,沿渭河谷(甘肃东部和陕西中西部)的人群表现出与高原人群更接近的遗传亲缘关系。相比之下,来自山西西南部、陕西东部和河南中西部的人群表现出更为同质的遗传特征,仅由中国黄河中新石器时期(China_YR_MN)血统代表。黄河下游地区主要以三种血统为特征:当地狩猎采集者相关血统、东亚南部相关血统和中国黄河中新石器时期相关血统。人类群体的遗传结构是由跨越地理、语言、文化和社会障碍的基因流动塑造的。71,72 古基因组数据使我们能够更好地理解人类基因流动与物质文化之间的复杂相互作用。73 在中新石器时期,黄河流域拥有明显不同的考古文化,这从工件组合和埋葬习俗的变化中可以看出。在黄河中上游流行的仰韶文化主要源自老官台文化和裴李岗文化,而黄河下游的大汶口文化则起源于北辛文化。74 在仰韶文化内部,独特的地方变体出现,表现出共性和差异。75,76 一些学者甚至将仰韶文化划分为不同的文化。77,78
我们的观察表明,文化因素在塑造黄河流域人口的遗传结构方面起着重要作用。然而,仅凭这些因素无法完全解释所观察到的遗传变异。地理因素在塑造遗传多样性方面也起着至关重要的作用。79,80,81,82 值得注意的是,地理距离在更细微的层面上影响遗传变异,并且可以独立于文化边界运作。82,83 例如,随着地理距离的增加,遗传相似性往往会减弱,这突显了一种文化分类无法完全捕捉的基因流动的渐进障碍。黄河流域包含不同的地理单元,其人口可分为三组:渭河组、陕西东部/山西西南部/河南组和下游黄河组。为了研究地理距离与遗传变异之间的关系,我们进行了皮尔逊相关性测试。我们的结果显示,地理距离与共享遗传漂变之间存在显著的负相关,这由外群f3结果所示(皮尔逊相关系数 = −0.412,p < 0.05,表S1F)。这表明地理因素在塑造黄河农民的人口结构方面起着关键作用。
粟农业和人口在中新石器时代的共同扩散
本研究考察了中新石器时期黄河流域及其邻近地区的人口遗传结构。我们的发现揭示了YR_MN祖源在整个黄河流域的显著分布,表现为多个方向的明显扩张:向北至河套地区(与庙子沟文化相关)、向东北至西辽河谷(与红山文化相关)、向南至长江中下游流域(与大溪文化、崧泽文化和良渚文化相连)、向西至青藏高原东北部(与马家窑文化相关)、向东至黄河下游(与大汶口文化相关)。这种基因扩散对周围社区产生了重大影响,与庙底沟文化的扩张和粟农业的兴起同步发生。
在7,000至5,000年前,粟作农业从几个可能的驯化中心向各个方向传播:向东至山东半岛和朝鲜84,85,86,向西至青藏高原东北部87,向南至长江中下游流域46,88,89,90,91,92,93,94,甚至到达东南沿海95。尽管在长江中下游地区发现了粟的遗存——如大水田遗址(6,031-5,933校正BP)和城头山遗址(5,800校正BP),以及安乐遗址(5,750校正BP)、上山遗址(6,500 BP)和青城墩遗址(6,000-5,300 BP)——但长江流域粟类作物消费的证据直到新石器时代晚期才出现96,97,98。因此,粟农业在中新石器时期在该地区的作用有限。
同时,庙底沟文化蓬勃发展,对邻近文化产生了重大影响。这种影响体现在红山、大汶口、大溪和崧泽文化中发现的具有庙底沟文化特征的器物和彩陶风格53,99,100。庙底沟文化的扩张促成了一个庞大文化共同体的形成,最终为夏、商、周王朝的出现奠定了基础53。总之,我们的研究强调了黄河农民及其粟农业在中新石器时期的共同扩散。
藏东高原与黄河上游在约5,800年前的基因互动
青藏高原,常被称为"地球第三极",平均海拔超过4,000米,是人类从非洲迁徙到具有挑战性的新栖息地的典型例子。68,101约500万藏族人已适应这种极端环境,并在高原上居住了数千年。然而,这些现代高原人口的起源仍然模糊不清。证据表明,早期人类在中部高原的存在可追溯到30,000年前,但这些早期狩猎采集者对当代藏族人的基因影响似乎有限。1012021年,Wang等人发现核心现代藏族人与古代尼泊尔人之间存在显著的基因相似性,揭示了与黄河农民的强烈基因联系。67这一发现表明,现代藏族人不太可能是古代藏族狩猎采集者的直系后代。此外,2023年的研究确定,现代高原祖先的主要来源可能起源于东亚北部,这种基因特征形成于5,100年前。68这些结果突显了藏族人口起源的复杂性,表明他们的遗传遗产相当大一部分来自北方平原的农业社区,而非高原早期的狩猎采集者。
我们的分析证实,与东亚北部其他地区相比,高原人口与黄河上游农民之间存在更强的基因关系。这通过基因亲和力与高原人口之间的显著负相关与经度(而与纬度无明显相关)得到证明。qpAdm模拟结果表明,黄河上游人口拥有19.5%至55%的高原相关祖先,平均为33.6%。值得注意的是,来自大地湾和圪垯川遗址的两个个体被发现携带超过50%的高原相关祖先。一个个体,Yangshao_DaDiWan_1,可追溯到约5,800校准BP,展示了与丹尼索瓦人相关的适应性EPAS1单倍型,标志着他们是已知最古老的现代人类,也是第一个被确认携带这种高海拔适应性特征的黄河农民。f4分析表明,DaDiWan_1与黄河农民人口比与高原人口共享更多的基因漂变。这两个结果都暗示,高原人口的主要基因祖先可能源自黄河上游地区的群体。
Yangshao_DaDiWan_1获得适应性EPAS1基因引发了有趣的问题。我们提出两个假设来解释这种基因亲和力:(1)从黄河中游迁移到上游的粟农可能通过适应当地环境和与周围人口交配,逐渐发展出独特的基因组结构,整合了高原生存的必要元素,如适应性EPAS1单倍型,从而使他们能够作为后续高原人口的直接祖先在高原上定居。(2)另一种可能是,拥有适应性EPAS1单倍型的高原人口可能迁移到黄河上游并与当地人口混合。虽然最终解释仍不明确,但这两个假设都表明高原与黄河上游人口之间的互动至少可追溯到5,800年前。此外,在江西沟2发现的陶片表明,青藏高原与低地文化之间的文化联系可追溯到7,000年前。102,103来自假设源区域的进一步基因组数据将对理解人类适应极端高原环境的更广泛影响至关重要。
黄河流域的南方血统:起源于长江流域,而非东南沿海
长江地区的基因组数据保存非常有限,导致该地区稻作农业人口的遗传结构基本上尚未被探索。为了弥补这一研究空白,我们对长江中下游地区的三个中新石器时代考古遗址进行了研究。我们的分析显示,这些核心长江遗址的个体表现出遗传同质性,与中国东南部旧石器时代晚期和新石器时代人群共享共同祖先,同时与黄河流域的农民在遗传上有明显区别。我们在黄河中下游地区的人口中发现了显著的南方祖先来源,表明南北方群体之间的互动超出了沿海地区。南方血统的引入对塑造黄河下游的遗传景观起到了至关重要的作用,那里的早期和中新石器时代北方沿海人口平均显示出超过30%的南方血统。这一点得到了基因亲缘性与中国南方人口之间存在显著正相关的支持,并且这种相关性与经度有关,而与纬度没有相应的相关性。我们的研究结果表明,黄河流域存在的南方血统很可能起源于地理上接近的长江流域,而非来自东南沿海的早期人群。这一发现挑战了之前认为沿海人口主要与中国东南部互动的假设。59,64
双向基因交流不仅发生在沿海地带,也出现在长江中游地区。在位于南阳盆地的朱岗遗址(黄河和长江文化的过渡区),我们检测到14.7%的南方血统,可追溯至长江流域。该地区不同的占据阶段反映了多样化的文化从属关系,突显了南北方人口之间的动态互动。这些发现意味着,稻作农业的扩张(可能由长江地区的人口迁移推动)可能早在中新石器时代就已开始,这修正了之前认为南方基因流向中原地区主要发生在随后的龙山时期的观点。62
台湾原南岛人的起源延伸至长江流域北部
南岛人的起源和扩散一直是学术界密切关注的课题。通过考古学、体质人类学、语言学和遗传学,学者们提出了几种理论,包括新几内亚理论、马来理论、台湾理论和大陆理论,其中后两种理论获得了最多的认可。104 考古证据表明南岛人与中国东南沿海地区存在联系。90,91 对岛屿东南亚的分析表明,南岛人群体拥有与台湾原住民更紧密相关的祖源。105 近期基于古DNA的遗传发现为这一争论增添了新的维度。2020年,研究人员发现大陆东南亚人群与台湾海峡岛屿人群表现出显著的遗传亲缘关系,支持了原南岛人起源于中国东南部的假说。59 随后2021年,Wang等人提出了一个假说,认为南岛人的遗传祖源可能有更北方的起源,潜在地可以追溯到长江流域的新石器时代农民。67 这些发现强调了在理解南岛人扩散时考虑中国南方起源的重要性。然而,由于缺乏直接的古DNA证据,这一假说尚未得到确认。
为了验证这一假说,我们系统地比较了新石器时代长江流域农民、大陆东南部人群和现代台湾南岛语族人群的遗传特征。104,106,107 我们的结果显示,现代台湾南岛语族人群与长江流域农民的遗传亲缘关系比与晚新石器时代之前的大陆东南部人群更为密切。现代台湾南岛语族人群可以被模拟为长江流域农民和当地台湾人群的混合。这一发现提供了初步证据,表明台湾原南岛人群的起源可能进一步延伸到长江流域北部,突显了新石器时代长江流域移民在塑造南岛人扩散中的作用。然而,要确定性地证实这一长江起源假说,还需要进一步的古DNA研究。
研究局限性
本研究面临几个局限性:(1) 来自长江流域的古基因组稀缺,限制了对稻作农业人口遗传结构的全面分析。这一限制可能阻碍了对新石器时期黄河和长江流域人口之间遗传交流的更深入理解。(2) 尽管在理解现代高地群体的人口历史方面取得了重要进展,但由于这些地区及其假设源区域缺乏更古老的基因组数据,他们迁移和适应高海拔环境的详细轨迹仍不清楚。(3) 考虑到混合图的重要性,我们还对混合图空间进行了全面探索,并确定了多个在统计上等效但代表生物学上不合理且不稳定情景的拓扑结构;因此,我们保守地将此类结果从文章中排除。未来在这些地区的采样工作,结合人口遗传分析方法的创新,可能会提供关于迁移和适应过程的确定性证据。在未来研究中解决这些差距对于更完整地重建东亚人口历史至关重要。
**长江流域样本
福泉山遗址**
福泉山遗址(FQS,31°09'32"N,121°07'27"E)位于上海市青浦县。自1979年以来,该遗址已多次进行发掘,出土了马家浜、崧泽和良渚文化的重要遗存。我们的样本取自1984年的考古工作,通过地层分析和从墓葬中提取的文物,被确认为属于晚期崧泽文化126。共发掘了17座墓葬,揭示了由鼎、豆、罐、壶和杯组成的基本陶器组合。此外,一些陶器上还刻有符号。渔猎工具数量减少,而农业工具呈现增长趋势。对墓葬层炭化物的放射性碳测定显示年代为公元前3644-3363年127。在这个遗址,我们成功测序了两个个体,其中一个样本通过放射性碳方法测定年代为公元前3513-3425年(T8M2)。
马桥遗址
马桥遗址(MQ,31°01'51"N,121°22'06"E)位于上海市闵行区马桥镇。该遗址于1960年和1966年进行了发掘。在第五层发现了10座墓葬;所有墓葬都是通过在平地上堆土建造而成,而非地下坑穴。这种埋葬方式在上海的广富林遗址和崧泽墓地也有发现。所有个体都呈仰身直肢位。在M4、M6和M7中发现了少量随葬品。该遗址发现了良渚文化的典型文物。我们成功测序了该遗址的一个个体,样本放射性碳测年为公元前3372-3330年。
大溪遗址
大溪遗址(DX,31°05'28"N,109°52'45"E)位于重庆市巫山县。该墓地自上世纪以来已多次发掘,揭示了与大溪文化相关的众多墓葬。大多数墓葬为竖穴墓,早期的特点是屈肢仰身埋葬,后期过渡为直肢仰身埋葬。在该遗址发现的大溪文化代表性文物包括桶形罐、深腹高圈足豆和曲腹杯。此外,该遗址的大溪文化还表现出仰韶文化的元素,如红黑彩陶52。该遗址还观察到将龟和鱼埋入墓葬的独特习俗。我们在该遗址产生了四个基因组数据集,其中两个样本通过放射性碳方法测定年代为公元前4257-4155年(M21)和公元前4172-4042年(M10)。
**黄河流域样本
刘林遗址**
刘林遗址(LL,34°21'08"N,118°00'49"E)位于江苏省邳县西北30公里处。1964年,共发掘了145座墓葬。这些墓葬根据出土的文物分为上下两层。埋葬方式包括直肢仰身埋葬、屈肢仰身或侧身埋葬,以及一些独特形式,如交叉腿/臂埋葬、折头葬和二次埋葬128。墓葬还根据埋葬个体数量进一步分类为单人和双人埋葬。刘林遗址是江苏北部的代表性遗址,被称为"刘林类型"。一些陶器展现出仰韶文化的特征。随葬品数量差异显著,从零到三十二不等,反映了社会经济分化。我们测序了三个样本:一个属于早期大汶口文化(缺失墓号-1,公元前4299-4253年),两个来自中期大汶口文化(M204,公元前3587-3529年)。
朱岗遗址
朱岗遗址(ZG,32°42'53"N,112°50'22"E)位于河南省南阳市。该遗址的所有样本都来自一个集体墓葬。我们测序了五个样本,其中两个直接放射性碳测年为公元前3950-3788年(朱岗①)和公元前3800-3696年(朱岗⑤)。
双槐树遗址
双槐树遗址(SHS,34°45'23"N,113°01'18"E)位于河南省巩义市双槐树村。该遗址由郑州市文物考古研究院于2013年至2020年进行发掘。该遗址特点是中心居住区被两个同心内沟和第三个外沟环绕。在内沟与外沟之间的夹沟区域内,发现了一个大型墓地,被确认为中原地区晚期仰韶文化时期(公元前3500-2900年)最大的墓地。已发掘超过1,600座墓葬,收集了287个个体129。炭化植物遗存表明,农业是早期居民的主要生产活动130,131。我们从该遗址的九个样本中产生了基因组数据,其中两个样本直接放射性碳测年为公元前3333-3214年(M469)和公元前3344-3093年(M498)。
孙庄遗址
孙庄遗址(SZ,34°45'09"N,113°36'48"E)位于河南省郑州市中原区孙庄村东南。考古发掘发现了从仰韶到龙山时期的多个文化层。共发掘了55座墓葬,年代从仰韶文化到宋代。大多数仰韶墓葬为竖穴墓,死者呈直肢仰身位。值得注意的是,孙庄个体的龋齿发生率较高,表明存在更复杂的农业经济132。我们从该遗址的三个个体中获取了基因组数据,其中两个样本直接放射性碳测年为公元前3242-3102年(M14)和公元前3119-3009年(M8)。
镇城遗址
镇城遗址(ZC,37°56'42"N,112°29'11"E)位于山西省太原市尖草坪区。2021年,山西省考古研究所和太原市文物考古研究所联合考古队的发掘发现了98个灰坑和两座仰韶文化房址,以及6座明清时期墓葬。出土了大量陶器、石器和动物骨骼。AMS-14C测年表明镇城遗址年代约为5042-4835 BP。在6号灰坑(H6)中发现了两具人骨,一名约28岁的成年男性和一名约2岁的幼年女性。对遗骸的同位素分析表明,饮食主要由C4植物和大量动物蛋白组成133。
辕村遗址
辕村遗址(YC,35°09'00"N,111°13'13"E)位于山西省运城市夏县。该遗址揭示了多个历史时期的遗存,包括晚期仰韶文化、庙底沟二期文化、龙山文化、二里头文化、二里冈文化以及汉代和宋代134。我们从4号灰坑(H4)获得了基因组数据,样本放射性碳测年为公元前2778-2621年。
南寨子遗址
南寨子遗址(NZZ,34°32'40"N,110°14'47"E)位于陕西省潼关县东南约0.6公里的台地上。2010年,陕西考古研究院对该遗址进行了考古调查和发掘。它标志着仰韶文化在关中地区的最东端延伸。在北部区域,发现了一个半坡文化墓地,包含四座墓葬135。南寨子出土的半坡文化文物与横镇和元君庙遗址的文物显示出很强的亲缘关系。共收集了18具人骨遗骸,代表了两性和所有年龄段。其中,M2墓葬包含六具人骨,包括两名成年男性、一名成年女性、一名儿童和一名性别不明的成年人136。我们成功测序了该遗址的一个个体,确认为男性,样本放射性碳测年为公元前4333-4227年。
吴中遗址
吴中遗址(WZ,34°22'42"N,109°12'50"E)位于陕西省西安市临潼区。2023年,陕西考古研究院对该遗址进行了发掘,发现了墓葬、灰坑、陶窑和各时期的房址基础。共发掘了14座墓葬,分布在聚落内外。其中,12座为竖穴墓,2座为房址墓,所有墓葬均无葬具,呈直肢仰身位。我们测序了四个样本,其中两个直接放射性碳测年为公元前4136-4054年(M112成人)和公元前4135-4055年(M111)。
东营遗址
东营遗址(DY,34°32'19"N,109°05'16"E)位于陕西省西安市高陵区,由陕西考古研究院于2016年至2017年发掘。该遗址出土了各个时期的考古遗存,包括仰韶文化史家类型、客省庄二期文化,以及战国至汉代和宋元时期。共出土19座墓葬,其中9座可归属于史家类型。这些史家类型墓葬特点是长方形竖穴,单人直肢仰身埋葬。大多数墓葬没有随葬品,除了M4,其中包含一个圜底钵。此外,从M11和M20中还发现了几件陶片。我们成功测序了东营遗址的两个个体,直接放射性碳测年为公元前3661-3625年(M4)和公元前2939-2881年(M20)。
灰坡岭遗址
灰坡岭遗址(HPL,34°45'37"N,109°10'45"E)位于陕西省渭南市富平县。2023年,陕西考古研究院对该遗址进行了发掘,发现了一条护城河、47个灰坑、7条灰沟和5座墓葬。大多数遗存属于新石器时代,特别是史家类型和庙底沟二期文化。我们从M8获得了基因组数据,放射性碳测年为公元前3591-3528年。
杨官寨遗址
杨官寨遗址(YGZ,34°32'19"N,109°05'16"E)位于陕西省西安市高陵区。自2004年以来,陕西考古研究院对该遗址进行了多次发掘。最重要的发现之一是属于庙底沟文化的护城河和墓地,这是中国首次确认的这一文化的大型墓地。迄今为止,已发掘超过500座墓葬,年代约为5,700至4,900年前137。墓葬分为三类:带侧室的竖穴墓、竖穴墓和带二次台阶的竖穴墓,以第一类最为普遍138。大多数墓葬不含随葬品,只有少数包含有限数量的陶器、骨器和贝器,以及玉器和石器。在一些情况下出土了颜料,在一个儿童墓葬中发现了一个完整的龟壳。我们从该遗址测序了九个样本,其中两个样本直接放射性碳测年为公元前3333-3214年(M353)和公元前3240-3103年(M449)。
马腾空遗址
马腾空遗址(MTK,34°12'42"N,109°02'15"E)位于陕西省西安市马腾空村,位于浐河西岸的第二级台地上。遗址面积约3公顷。2016年至2018年,陕西考古研究院对该遗址进行了系统发掘。遗存跨越五个时期,包括仰韶时期、东周、隋代、明代和清代。在居住结构周围出土了34座晚期仰韶时期的墓葬。这些墓葬特点是尸体呈直肢仰身位,头朝南脚朝北。这些墓葬中没有发现陶器,只有少量石器。值得注意的是,M1257墓葬中包含一把石斧和一把石铲139。我们测序了来自M1257的样本,放射性碳测年为公元前3343-3097年。
王阳畔遗址
王阳畔遗址(WYP,37°59'31"N,109°17'27"E)位于陕西省榆林市横山县城关镇马家毛村王阳畔组,于2010年发掘。该遗址主要特点是晚期仰韶时期的遗存,同时有一些早期龙山时期的文物。考古学家发现了七座房址基础、62个灰坑和三座墓葬。墓葬为东西向长方形竖穴墓,尸体呈直肢仰身位,没有发现随葬品。植硅体分析表明,居民在晚期仰韶文化时期主要种植粟140。我们的样本采集自1号灰坑(H1),测序了一个个体。样本放射性碳测年为公元前2816-2668年。
圪垯川遗址
圪垯川遗址(GDC,35°00'20"N,106°12'19"E)位于甘肃省天水市张家川县。该遗址于2021年发掘,跨越多个时期,包括仰韶文化、齐家文化和历史时期。值得注意的是,仰韶时期的遗存覆盖约160,000平方米的区域,可分为三个阶段:半坡期、庙底沟期和晚期仰韶文化。据推测,该遗址有功能区划,如居住区、陶器生产区和墓葬区。对人类以及猪和狗的碳同位素分析表明,圪垯川居民主要依赖C4食物44。我们从圪垯川遗址测序了九个样本,直接放射性碳测年为公元前4300-3500年。
王家阴洼遗址
王家阴洼遗址(WJYW,34°52'28"N,105°40'38"E)位于甘肃省天水市秦安县东北部。该遗址于1981年由甘肃省博物馆大地湾发掘队发掘。共出土63座早期仰韶文化墓葬。这些墓葬分为东西两部分,西部的死者面向东南,东部的面向北东北。墓葬包括单人、多人、二次和罐棺葬,大多数在骸骨左侧设有额外的椭圆形竖穴用于放置随葬品。布局表明墓地作为公共氏族埋葬地。王家阴洼遗址的遗存与关中地区的史家类型有相似之处141。我们从该遗址测序了八个样本,其中三个样本直接放射性碳测年为公元前3773-3650年(M36)、公元前3951-3794年(M57)和公元前3950-3776年(QW-缺失墓号)。
大地湾遗址
大地湾遗址(DDW,34°52'28"N,105°40'38"E)位于甘肃省天水市秦安县东北部的少店村。该遗址占据历史跨度从7800到4800 cal BP,是理解仰韶文化迁移和中国西北部粟作农业发展的关键遗址。大地湾进行了广泛的考古工作,其占据历史分为五个文化阶段,从前仰韶到常山文化:I期(7,800-7,300 cal BP,前仰韶)、II期(6,500-5,900 cal BP,早期仰韶)、III期(5,900-5,500 cal BP,中期仰韶)、IV期(5,500-4,900 cal BP,晚期仰韶)和V期(4,900-4,800 cal BP,常山文化)142。尽管农业实践始于I期,但当时还处于起步阶段,以狩猎采集经济为主143。在II期至IV期发现了粟和黍,表明粟作农业已发展到一定程度144。我们从该遗址产生了两个基因组数据,均直接放射性碳测年为公元前3955-3783年(GDT4③)和公元前3951-3764年(GDT4③)。
山那树扎遗址
山那树扎遗址(SNSZ,34°27'20"N,104°02'07"E)位于甘肃省岷县。2012年和2013年,甘肃省文物考古研究所进行了两次发掘,覆盖面积约1,400平方米。这些发掘发现了196个遗迹,包括房址和储藏坑。文物表明该遗址属于早期马家窑文化,同时保留了中晚期仰韶文化的元素87。分析样本采集自2013年的发掘。两个样本经测序和直接放射性碳测年为公元前3248-3100年(T0210)和公元前3344-3093年(T0208H175)145。
这篇总结下:
- 仰韶老农兜里揣着小米 赢麻了, 向西到青藏高原(发现了最早的仰韶古人 5800年 且携带丹人高原基因),向北上西辽河,影响东北平原,向南与长江流域人群产生双向交流.
2.发现 Daxi_DaXi_2 与核心长江流域群体分离,拥有11.5% 更高的北方祖先成分,额外有5% 深度的青藏/中国西南 成分
3.补充了台湾原始南岛群体不仅与中国东南沿海有关,更与长江流域有关
Grock4总结:
文章详细总结:《东亚中石器时代小米和稻米农业人口的基因组历史》
这篇发表于 Cell Genomics 杂志的文章(在线发布于2025年8月19日)对黄河流域和长江流域中石器时代人口进行了详细的基因组分析,这些地区分别以小米和水稻的驯化为世所知。该研究通过分析这些地区的基因历史,揭示了农业扩张对人类遗传的影响。以下是对文章的详细总结,包括引言、方法、结果、讨论和结论部分,涵盖基因分析、人口互动以及对人类历史的含义。
引言
引言部分强调,黄河流域和长江流域是中国乃至世界上最古老的独立农业中心之一,黄河流域以小米驯化为特征,长江流域则以水稻驯化为特征。然而,这些地区在中石器时代(Middle Neolithic)的遗传历史仍知之甚少。该研究旨在通过基因组数据分析,探讨遗传分化、基因流动以及农业扩张对人类遗传历史的影响。
方法
文章未在提供的内容中明确详细描述方法部分,但推断涉及从黄河流域和长江流域的74个中石器时代遗传样本中生成全基因组数据。分析可能包括古DNA技术,用于考察遗传变异、人口结构和基因流动的具体方法(如测序和统计分析)未在摘要中详述。
结果
结果部分揭示了多项关键发现:
- 中石器时代黄河流域农民表现出明显的遗传子结构,这可能源于小米农业在中石器时代扩张期间与周边群体的互动。
- 黄河流域和长江流域中石器时代人口之间存在双向基因交流,支持混合农业的“人口扩散模型”(demic diffusion model),即农业传播涉及人口迁移和文化交流。
- 上游黄河流域人口与青藏高原人口在遗传上相关联,并携带有史以来最早的适应性EPAS1单倍型(约5,800年前),这是现代人类中最早的高海拔适应变异。
- 长江流域水稻农民显示出与新石器时代至今中国东南沿海地区以及南岛语系人口的遗传亲缘关系,该研究将原南岛人的遗传起源追溯到更北部的长江盆地。
讨论
讨论部分将这些发现置于中石器时代农业扩张及其对人类遗传多样性的影响的背景下进行解读。黄河流域的独特遗传子结构表明与邻近群体的复杂互动,可能由小米农业的传播驱动。 黄河和长江之间双向基因流动表明农业系统相互连接,并涉及人口移动。 上游黄河流域个体中早期EPAS1单倍型的存在突显了早期的遗传适应,可能与环境压力相关。 长江流域人口与南岛语系的遗传亲缘关系表明原南岛人的起源位于长江盆地,这重塑了对他们扩散和文化历史的理解。
结论
结论部分强调,这些发现为东亚中石器时代人口的基因组历史提供了新洞见,特别是中石器时代农业扩张如何塑造了人类遗传多样性。该研究突出了黄河流域和长江流域在理解早期农业社会、人口互动以及现代东亚和南岛人口遗传基础方面的关键作用,并为进一步研究古代人口动态及其现代遗产奠定基础。
基因分析、人口互动以及对人类历史的含义
基因分析显示,黄河流域和长江流域人口之间存在显著分化,黄河流域群体表现出与小米农业扩张相关的子结构,并与周边群体(包括青藏高原人口)互动。 双向基因流动支持人口扩散模型,表明农业传播涉及迁移和文化交流。 上游黄河流域个体中约5,800年前的EPAS1单倍型表明适应性遗传变化,可能与高海拔环境相关。 对于人口互动,该研究突出了长江流域水稻农民与南岛语系人口的联系,将原南岛人的起源追溯到长江盆地,这对理解他们的扩散和文化发展具有重要含义。 这些发现整体上提升了对中石器时代农业扩张如何影响东亚遗传多样性和人类历史的认识。
 陕公网安备61011302002223号
陕公网安备61011302002223号