古代基因组揭示了玛雅古典时期的人口变迁轨迹
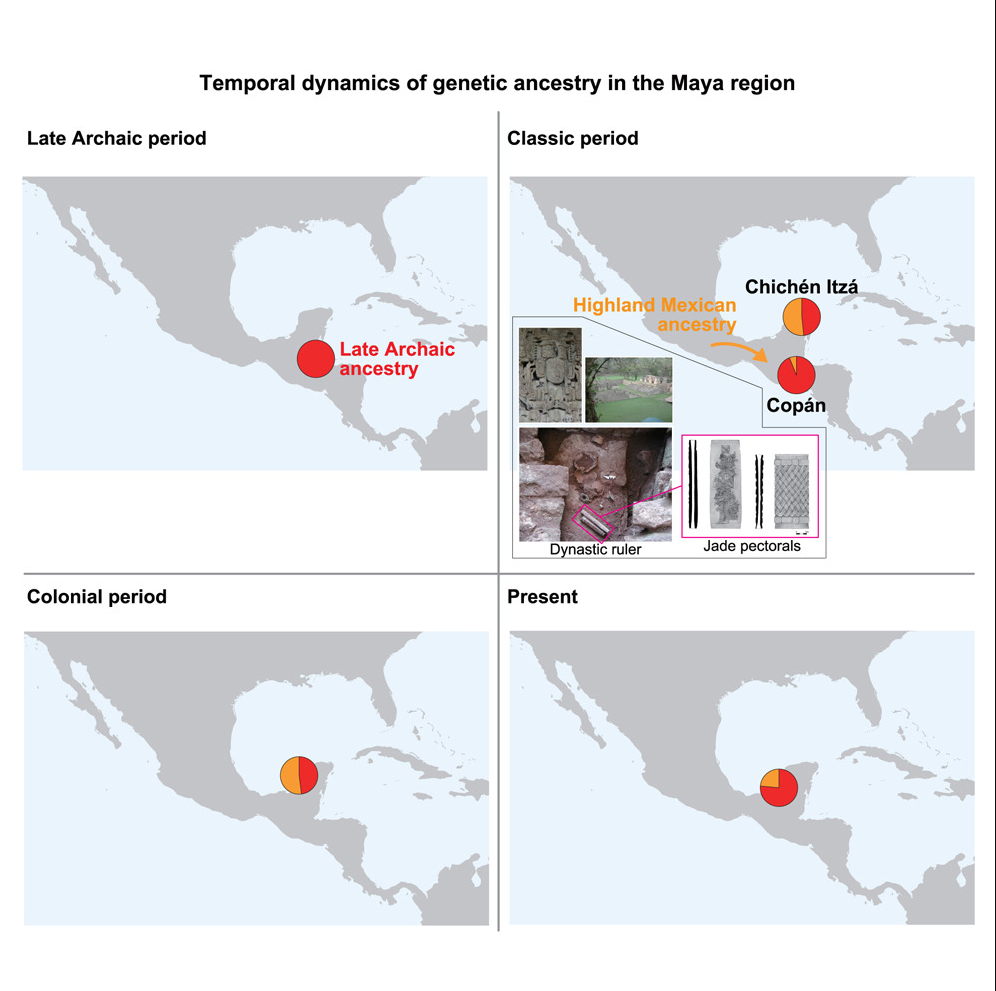
## 要点 - 从科潘遗址的七具个体中提取了古代基因组 - 从晚期原古时期开始,玛雅地区保持着本地血统 - 在早期古典时期发现了流入科潘的基因流动 - 基因证据显示人口下降与文明衰落同步发生 ## 概要 科潘是古典玛雅文明最东南端的一个主要都城,是连接中美洲和南美洲的十字路口。在公元426/427年,该城市建立了一个持续约400年的王朝。尽管有大量的历史和考古记录,但关于科潘居民的基因特征仍然了解有限。 本研究展示了来自7个古典时期科潘个体的基因组,其中包括一位可能的王室成员及其陪葬牺牲者。我们的分析表明,与古典科潘文化相关的人群与伯利兹晚期原古时期人群、墨西哥奇琴伊察的晚期古典玛雅人、坎佩切
要点
- 从科潘遗址的七具个体中提取了古代基因组
- 从晚期原古时期开始,玛雅地区保持着本地血统
- 在早期古典时期发现了流入科潘的基因流动
- 基因证据显示人口下降与文明衰落同步发生
概要
科潘是古典玛雅文明最东南端的一个主要都城,是连接中美洲和南美洲的十字路口。在公元426/427年,该城市建立了一个持续约400年的王朝。尽管有大量的历史和考古记录,但关于科潘居民的基因特征仍然了解有限。
本研究展示了来自7个古典时期科潘个体的基因组,其中包括一位可能的王室成员及其陪葬牺牲者。我们的分析表明,与古典科潘文化相关的人群与伯利兹晚期原古时期人群、墨西哥奇琴伊察的晚期古典玛雅人、坎佩切的殖民时期玛雅人以及现代玛雅人形成一个基因聚类。
以伯利兹晚期原古时期人群作为本地血统的来源,我们发现从早期到中期古典时期开始出现与墨西哥高地人群相关的基因流入,突显了基因流动在古典玛雅国家形成过程中的作用。有效人口规模的估计表明,在古典时期末期,当古典玛雅文明经历广泛的动荡和崩溃时,人口出现了下降。
关键词
引言
科潘是古典玛雅文明(公元250-950年)最重要的中心之一。科潘最早的人类居住可以追溯到早期前古典时期(公元前1000年以前)。在这个时期,位于洪都拉斯西部靠近危地马拉边境的科潘山谷出现了以玉米种植为主的小型农业社区。
早期古典时期(公元300-400年)开始出现玛雅建筑和碑文等纪念性建筑。根据目前的碑文解读结果,一个王朝于公元426/427年建立。这个王朝先后有16位统治者统治这座古城及其政体。科潘作为政治、经济和祭祀中心蓬勃发展,考古学家在此发现了精英和平民的居住建筑。尽管人口估计在鼎盛时期达到约3万人,但这个政权最终在晚期古典时期末(约公元820年)崩溃。
关于科潘王国形成的一个长期假说认为,在非玛雅人居住的地区存在着一个玛雅文化飞地。据推测,属于广泛分布的玛雅精英网络的个体迁移到该地区,可能来自危地马拉中部佩滕地区,特别是蒂卡尔地区,他们与当地非玛雅居民通婚。考古证据表明科潘与其他中美洲城市,如蒂卡尔、危地马拉山谷的卡米纳尔胡尤和墨西哥山谷的特奥蒂瓦坎之间存在广泛的贸易和政治联系。
科潘的碑文揭示,第一位王朝创建者K'inich Yax K'uk' Mo'是一位外来者,他于公元426年掌权,427年抵达科潘。牙釉质同位素分析进一步证实了人口流动的广泛性,在科潘发现的人类遗骸中揭示了原居地的显著差异。值得注意的是,这种差异在精英墓葬中更为显著,包括第一位和后续国王。由于牙釉质的化学成分反映了婴儿期和幼年期消耗的食物和液体,这表明这些精英在迁移到科潘之前是在城外长大的,最终在此安葬。然而,同位素分析无法识别第一代移民之后的非本地个体。需要基因数据来探索本地和非本地人群之间的长期互动,以更好地理解随后的社会和文化变迁。
玛雅地区古基因组的先前分析揭示了一种可追溯到全新世早期(约9,300-7,300年前)的独特基因祖先,这可能是通过美洲南向迁移初期分化出的一支血统。到晚期古代时期(约5,600-3,700年前),该地区出现了来自更南方的基因流动。尽管后来的墨西哥高地血统(由瓦哈卡、米赫和萨波特克等土著群体代表)影响了现代玛雅人的基因构成,但其到达的具体时间仍不清楚。最近对尤卡坦奇琴伊察晚期古典时期基因组的分析提供了与该地区当代人群基因连续性的证据。
在本研究中,我们报告了七个新测序的科潘古基因组,均来自古典玛雅时期。其中包括一个个体(CpM13),他被埋葬在具有王室墓穴特征但位于卫城之外的墓中,随葬品表明他属于统治王朝。CpM13还具有最高的财富等级评分"5",这是基于埋葬形式、结构和内容的社会地位大致衡量。另一个测序个体(CpM12)被发现在这位精英个体附近,处于与祭祀埋葬相关的位置,其财富等级为"0"。通过将这些科潘基因组与涵盖南美洲、中美洲、北美洲和西伯利亚的更广泛的古代和现代基因组数据集相结合,我们的研究旨在加深对古典科潘人群的基因祖先和人口统计的理解,揭示玛雅地区东南边缘的人口流动和社会网络动态。
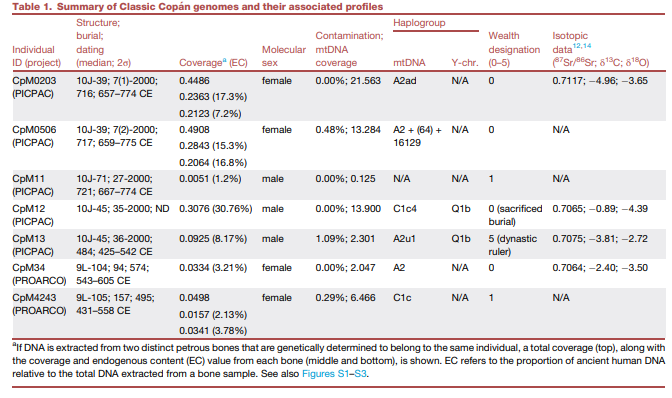
表1. 科潘古典时期基因组及其相关特征概述
注:如果DNA从两个不同的岩骨中提取,且这些岩骨经基因鉴定属于同一个体,则显示总覆盖度(上)以及每个骨骼的覆盖度和内源DNA含量(EC)值(中和下)。EC指骨骼样本中古代人类DNA相对于总DNA的比例。另见图S1-S3。
研究结果
科潘古代基因组
我们最初的筛查聚焦于从科潘遗址出土的16具古代骨骸,这些遗骸是作为两个项目的一部分被发掘出来的(科潘考古公园综合保护计划PICPAC和科潘考古项目PROARCO),这两个项目都由中村诚一领导(参见数据S1)。在这些个体中,有七具的内源人类DNA含量超过1%(表1),随后对这些样本进行了更深度的测序,覆盖度范围从0.033×到0.491×不等。这七个个体与五个不同的建筑结构相关(PICPAC:10J-39、10J-45和10J-71;PROARCO:9L-104和9L-105)。放射性碳测年显示所有个体都属于古典玛雅时期,并可分为两个时间组:早期至中期(约公元250-550年)或中期至晚期(约公元550-830年)古典玛雅阶段。
我们确认所有新测序的基因组都表现出死后损伤模式(图S1)和DNA片段长度较短(图S2),中位长度在37到42bp之间,并且现代人DNA污染水平较低(<1.09%)(表1)。亲缘关系系数估计显示两对个体之间可能存在二级亲属关系:CpM11和CpM34、CpM11和CpM4243(图S3)。然而,由于CpM11的覆盖度较低,这些亲缘关系估计可能不太可靠,因此我们在后续分析中排除了这个个体。其余六个个体的单倍体群属于A2或C1谱系,这两个谱系都被归类为泛美洲线粒体DNA单倍体群,在当今美洲各地的人群中普遍存在。
分子性别鉴定识别出三名男性,包括一个可能的祭祀埋葬者(CpM12),这个个体此前根据形态特征被认为是女性。两名男性CpM13和CpM12属于Y染色体谱系Q1b,这是当今美洲人群中最普遍的单倍体群,而另一名男性CpM11由于序列覆盖度不足,无法确定Y染色体单倍体群。为了在更广泛的美洲基因组景观中理解我们的数据,我们将古典科潘基因组与先前发表的古代个体的基因组数据(共709个古代基因组;图S4;表S1)以及现代人群数据进行了组合分析。
古典科潘的人群结构
我们基于外群f3测量值,通过多维尺度分析(MDS)探索了古典科潘与北美、中美和南美其他古代人群以及现代人群之间的全基因组常染色体亲缘关系。这项分析按照地理来源将古代人群分开,并识别出与加利福尼亚、加勒比(特别是原古时期)和安第斯山脉相关的不同聚类(图1A)。所有来自中美洲的人群——包括古典科潘、伯利兹早期全新世和晚期原古时期基因组、墨西哥古典和殖民时期玛雅基因组、巴拿马前西班牙时期基因组以及现代人群——倾向于聚集在一起(图1A)。值得注意的是,无论选择哪个维度,现代玛雅人都始终显示出与古典科潘最接近的基因亲缘关系(图S5)。来自其他地理区域的人群,如巴西、阿根廷和陶器时期加勒比地区的人群,在MDS分析的不同维度上都靠近或与中美洲聚类重叠(图S5),这表明这些人群之间可能存在基因联系。
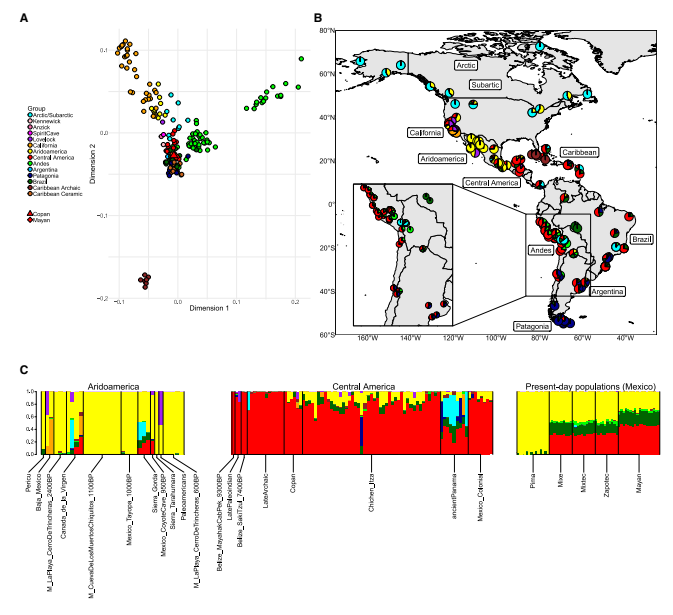
图1 在古代和现代美洲人群背景下的古典科潘基因组特征
这种共同祖先关系在我们的ADMIXTURE分析(K = 9)中得到进一步支持(图1B、1C和S6-S8)。其中一个成分(红色)在多个人群中普遍存在,包括来自中美洲、加勒比、安第斯山脉、巴西和阿根廷的人群(图1B)。这一祖先成分在中美洲个体中始终保持较高水平,特别是在玛雅地区的早期全新世(9,300-7,400年前)和晚期原古时期(5,600-3,700年前)个体中达到最高(图1C)。相比之下,后期玛雅人群,如古典科潘、奇琴伊察和墨西哥殖民时期,倾向于表现出另一个成分(黄色),这个成分在干旱美洲和现代皮马人群中占主导地位(图1C)。其他祖先成分主要反映地理分布(图S7):青色代表北极和巴西的一些早期全新世个体,橙色代表海峡群岛,紫色代表加利福尼亚其他地区,绿色代表安第斯山脉,深蓝色代表巴塔哥尼亚,棕色代表原古时期加勒比地区。
与玛雅地区人群的基因亲缘关系
我们通过计算f4统计量(形式为f4(姆布蒂人,X;Y,科潘))研究了古典科潘基因组与西伯利亚和美洲各地人群(包括188个古代和12个现代人群)之间的基因关系。这项分析将这200个人群中的每一个作为X进行迭代,其余人群被指定为Y。然后,我们根据一个人群相对于其他199个人群显示出与古典科潘更大基因亲缘关系的频率进行排名,Z分数>3.0(图2A;表S2;数据S2)。这项分析揭示了古典科潘人群与其他中美洲人之间强烈的基因亲缘关系。特别是,现代玛雅人排名第一,其次是墨西哥奇琴伊察的晚期古典玛雅人排名第二。
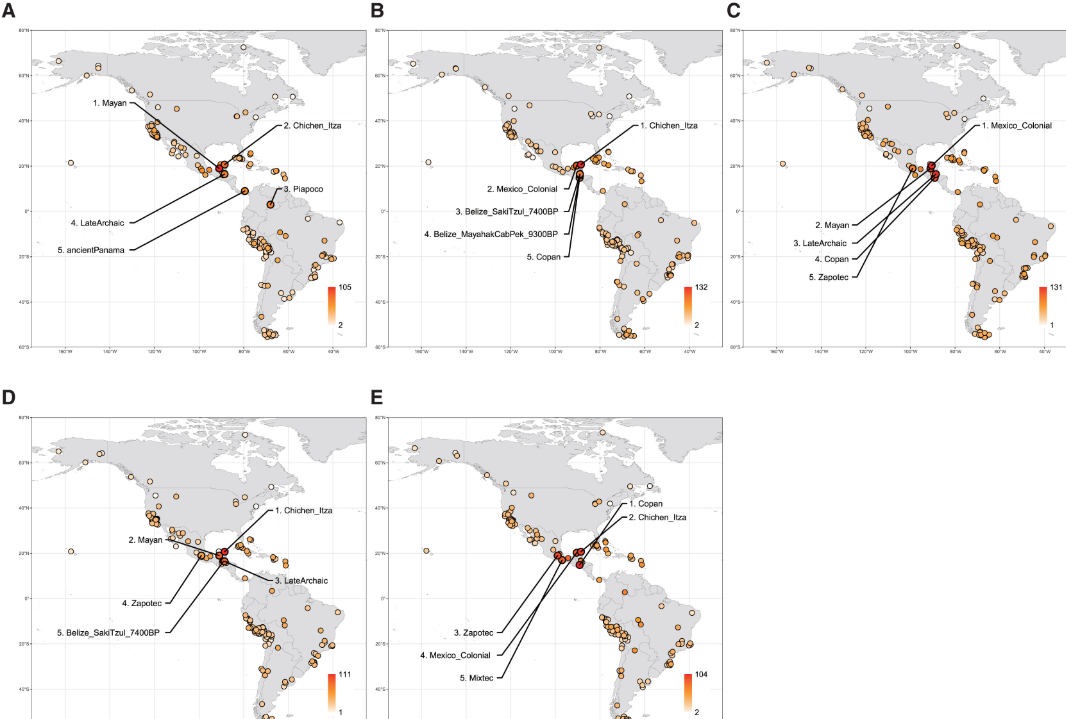
然后,我们通过将科潘分别替换为晚期原古时期、奇琴伊察、墨西哥殖民时期或玛雅人进行f4检验,探讨了这些基因亲缘关系是否在玛雅地区的古代基因组时间序列中得到相互支持(图2B-2E;表S2;数据S2)。这些额外的分析表明,相对于被检测的人群,排名最高的人群始终是较早或较晚的玛雅群体,这为这些人群之间随时间推移的基因联系提供了有力证据。特别是,正如之前研究报告的那样,伯利兹的早期全新世人群与该地区晚期原古时期人群表现出显著的基因亲缘关系。
基于refinedIBD使用插补数据进行的同源(IBD)分析进一步支持了玛雅地区人群之间的紧密时间联系(图S9和S10),随后进行了基于网络的社群检测。本研究中评估的插补性能与之前一项纳入更多古代美洲基因组的研究结果一致。来自晚期原古时期和古典科潘的所有个体,以及来自奇琴伊察和殖民时期墨西哥的大多数个体都被归类在同一个社群("1-1")中(图3和S11),这是一个在空间和时间上都具有多样性的社群"1"的一部分(图S12和S13)。这种聚类主要反映了地理和文化背景(图S12),与ADMIXTURE分析结果一致(图1)。总的来说,这些结果表明玛雅地区从晚期原古时期直到现在保持着一定程度的基因连续性。
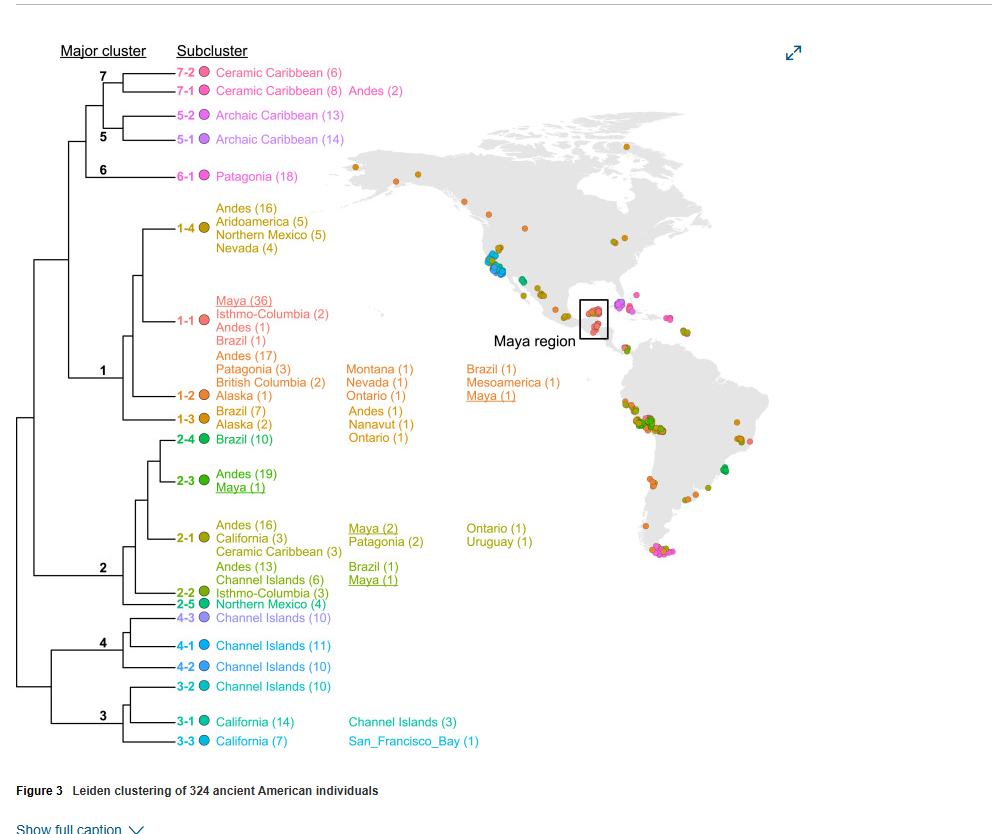
图3 324位古代美洲个体的Leiden聚类分析
古代和现代玛雅人群的遗传祖源
为了探索玛雅地区随时间推移的基因流动模式,我们直接比较了该地区不同人群配对(即晚期原古时期、科潘、奇琴伊察、墨西哥殖民时期和玛雅人)与所有其他美洲人群的基因亲缘关系(表2和S3),使用f4统计量,形式为f4(姆布蒂人,X;Y,Z),其中Y和Z代表一对玛雅人群,而X是我们数据集中的任何其他人群。这些f4检验表明,与晚期原古时期人群相比,后期玛雅人群始终与来自塞拉戈尔达(干旱美洲和中美洲之间的过渡地区)的前西班牙时期人群,或与来自瓦哈卡的墨西哥高地人群(包括米赫人、萨波特克人和米斯特克人)表现出显著的亲缘关系。相比之下,一些早期全新世个体,无论是来自伯利兹还是智利和阿根廷的,都与晚期原古时期人群表现出比后期玛雅人群更强的亲缘关系。
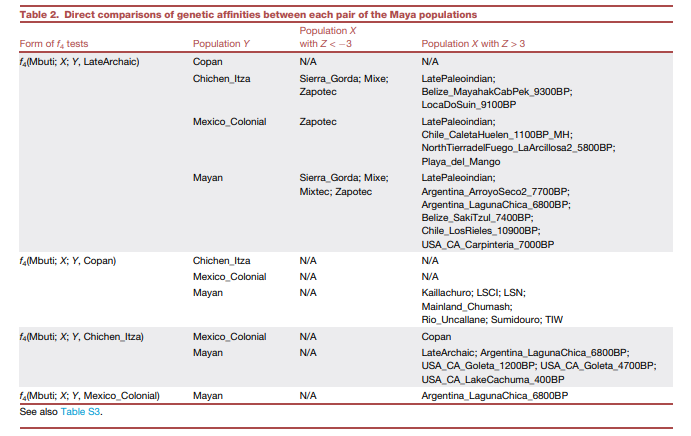
表2 玛雅各个人群之间基因亲缘关系的直接比较
我们使用晚期原古时期作为玛雅地区本地血统的来源,评估了从塞拉戈尔达或瓦哈卡流入古典时期、殖民时期和现代玛雅地区人群的基因(表S4)。我们的混合模型显示,除了古典科潘外,塞拉戈尔达不太可能是基因流动的来源,因为在玛雅人群中尾部概率低于5%。相比之下,当使用萨波特克人(一个始终与后期玛雅人群表现出基因亲缘关系的人群,表2)作为墨西哥高地血统的来源时,混合模型得到了所有人群的有力支持(图4)。
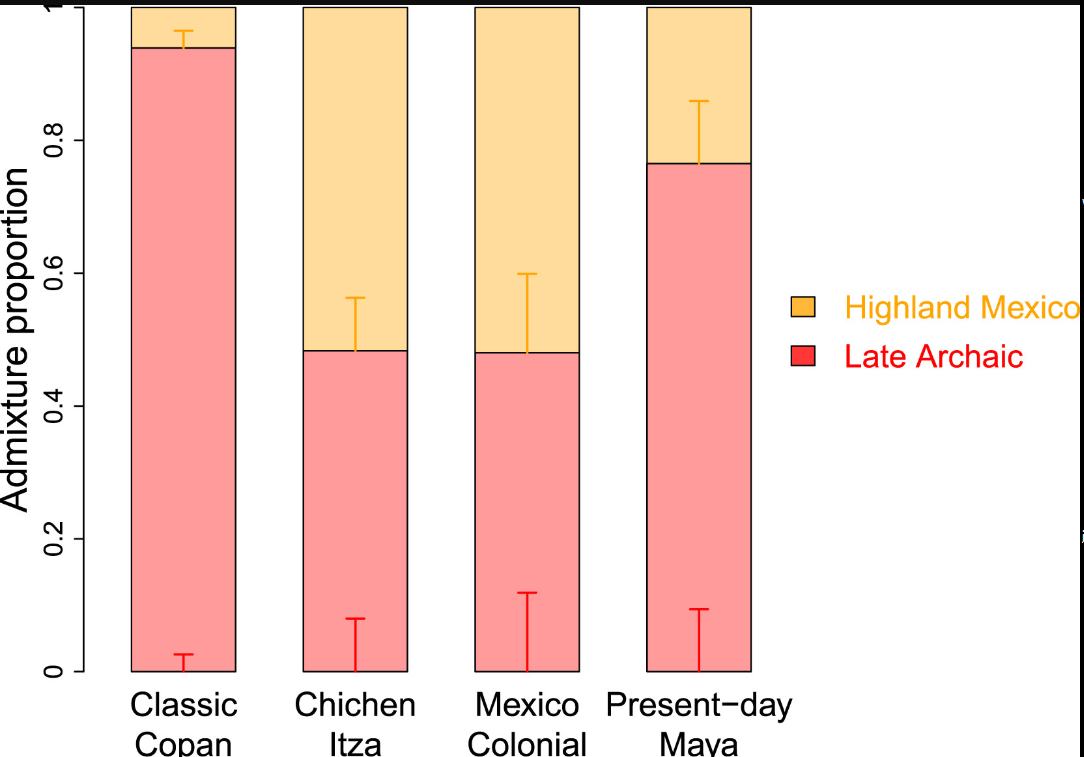
在古典科潘,晚期原古时期血统占主导地位,墨西哥高地血统仅占6.1%±2.6%。这种模式在不同时期群体和古典科潘个体中都保持一致。相比之下,来自奇琴伊察的晚期古典时期人群显示两种祖源的贡献几乎相等。这些比例在坎佩切的殖民时期人群中似乎保持稳定,但在现代玛雅人中又发生了转变,其中晚期原古时期血统占76.5%±9.4%。然而,考虑到奇琴伊察和殖民时期墨西哥人群中观察到的个体间显著差异,这种转变可能反映的是小样本量(即两个个体)的影响,而不是整个人群的趋势。
使用DATES方法,我们估计奇琴伊察人群中晚期原古时期和墨西哥高地血统之间的混合发生在497±159年前。考虑到奇琴伊察个体报告的校准日期中位数平均值为公元892年,这一估计支持墨西哥高地血统在早期到中期古典时期开始出现在玛雅地区。
玛雅地区人口崩溃的基因痕迹
我们利用IBD片段共享来推断人口规模的时间变化。平均IBD总长度的模式在不同时期和地区之间存在差异,这可能反映了人口统计学的差异。为了探索导致这种观察到的变异的历史因素,我们采用滑动窗口方法,使用每个时间窗口内个体之间的IBD共享作为IBDNe的输入,在特定时间点估计人口规模。
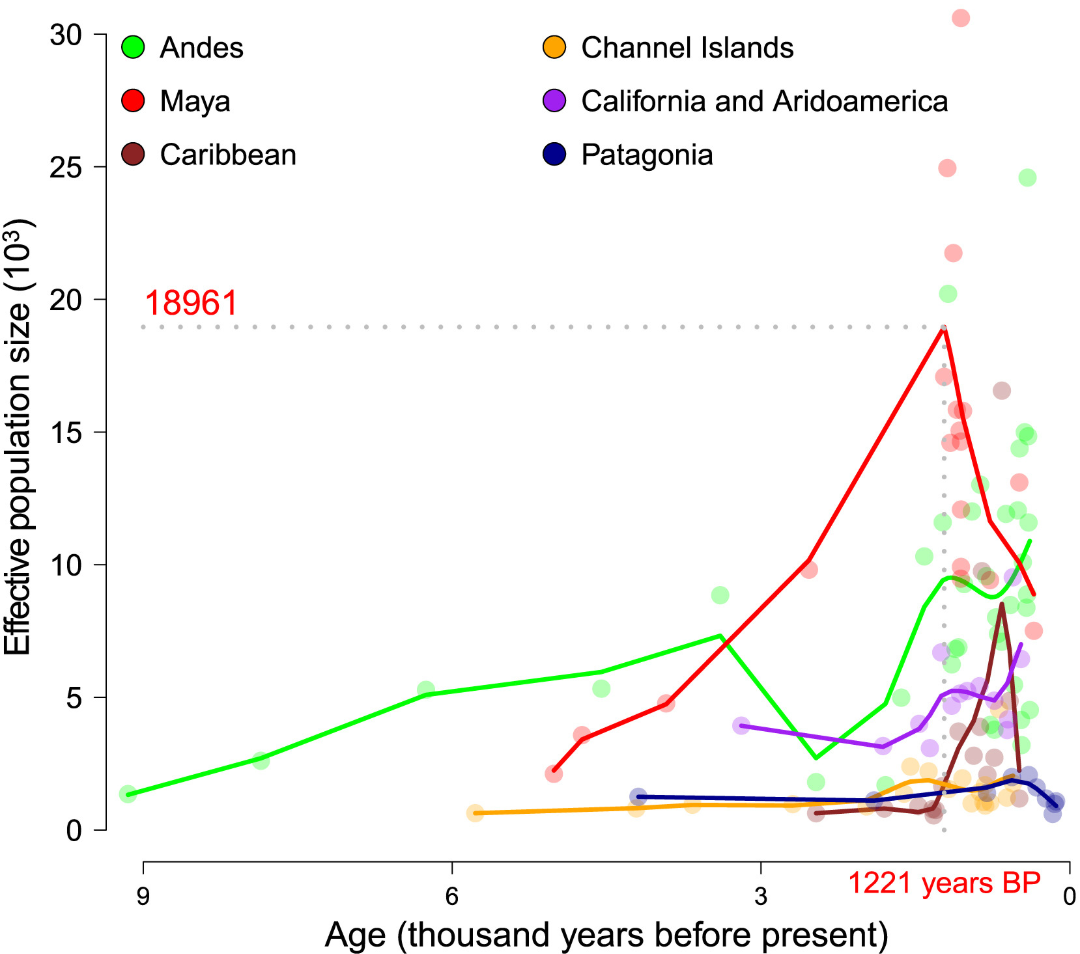
图5 美洲不同地理人群的人口规模随时间的变化
玛雅地区的人口规模似乎经历了显著增长,在大约1,221年前达到约19,000人的有效人口规模。这种增长可能与古典时期来自墨西哥高地的基因流动有关,同时也与农业的出现有关。这一点得到了古典科潘个体的支持,他们显示出对玉米作为主要食物来源的高度依赖(图S17)。这种增长在分析的六个地区中明显突出。例如,在海峡群岛或巴塔哥尼亚,人口规模随时间推移一直保持较小。加勒比地区近期的人口规模增长可能反映了从原古时期到陶器时期的基因和文化转变,这一点从其IBD模式中可以看出(图S16)。然而,玛雅地区的人口增长似乎在约1,200年前开始逆转,这与古典玛雅文明崩溃的开始时间相吻合。在安第斯地区也可以观察到同一时期的人口下降。不过,安第斯地区的人口规模在后期又回升,这可能与印加文明的兴起和扩张有关。
讨论
我们对新产生的古典科潘基因组与已发表的古代和现代美洲基因组的综合分析,揭示了古典玛雅时期的基因和人口变化。MDS、ADMIXTURE、f4统计和IBD的人口结构分析(图1、2和3)强有力地支持玛雅地区从晚期原古时期到现在的基因连续性。然而,这种连续性并非是排他的,因为该地区的时序基因组揭示了与墨西哥高地人群(可能来自瓦哈卡等地区)相关的基因祖源在早期到中期古典时期开始出现。
我们的分析表明,这种基因流动延伸到了位于古典玛雅国家地理边缘的科潘(图4),但相对于更北部的人群(如奇琴伊察)来说,这种基因流动较为稀释。这些发现突显了流动性(可能与玛雅精英网络有关)在科潘王国形成过程中的作用。从晚期原古时期到现在的强烈基因联系支持了考古研究提出的观点:本地人群与来自更大玛雅网络的成员的融合是玛雅政体扩张的一种策略。
从IBD片段共享推断的人口统计学表明,玛雅人口的有效规模在约1,200年前急剧下降(图5),这与古典时期的结束相吻合。这一时期还出现了频繁的严重干旱,发生在公元9世纪至11世纪之间,这被认为是导致内战、社会不稳定和古典玛雅文明崩溃的几个因素之一。需要注意的是,虽然这些干旱的时间和强度以及社会崩溃的过程在空间上存在变异,但我们的分析并未考虑这种变异性。因此,人口规模的估计反映的是一组古代玛雅个体的整体趋势,而不是代表任何特定的文化或地理背景。
总之,我们的研究提供了一个新的视角,表明玛雅地区本地血统的持久存在,同时也突显了超出传统玛雅领土边界的更广泛流动性。
研究局限性
这项分析存在一些注意事项。首先,我们的研究受到样本量小的限制,只涉及七个古典科潘个体,其中大多数基因组覆盖率较低。这些是统计检验和混合建模中众所周知的限制因素。其次,需要注意的是,个体的财富指定分数与其考古背景之间存在差异。财富指定测量基于之前研究概述的标准,这些标准可能并不完全适用于古代科潘社会的埋葬背景。事实上,对与玛雅文化相关的丧葬背景的深入分析显示,近90%被归类为低等级(即"0"或"1"分),只有5%被分配为"4"或"5"分(即高等级),这表明大多数人口都属于低等级类别。最后,我们的取样是非随机的,集中在来自相同住宅或庭院群体的个体(表1)。考虑到科潘遗址以及其他古典玛雅国家之间观察到的考古和同位素变异,更广泛和密集的古代基因组采样将对追踪国家内部人口的时间和区域多样性至关重要,这将为理解古典玛雅复杂人类社会的兴衰提供更全面的视角。
 陕公网安备61011302002223号
陕公网安备61011302002223号