古DNA揭示鲜卑联盟的起源和迁徙模式
鲜卑是匈奴帝国崩溃后东欧亚大陆最强大的游牧群体之一。然而,由于缺乏第一手的文字记录,鲜卑的起源及其与周边人群的关系仍然存在谜团。在此,我们对来自中国北方的9位鲜卑个体(约公元200年至300年)产生了基因组数据。通过结合文献中现有的基因组,我们建立了一个数据库,涵盖了几乎整个鲜卑时期以及他们之前和之后的样本,使我们能够将鲜卑置于时间背景中进行研究。 我们的研究决定性地解决了一个长期存在的假设,支持鲜卑起源于黑龙江地区,更具体地说是来自中国东北部大兴安岭山脉周边地区。我们还提供了直接的遗传证据,表明在他们最初向中原地区南迁的过程中,鲜卑仅从他们遇到的当地人群中接受了有限的外源基因贡献,但在定居
摘要
鲜卑是匈奴帝国崩溃后东欧亚大陆最强大的游牧群体之一。然而,由于缺乏第一手的文字记录,鲜卑的起源及其与周边人群的关系仍然存在谜团。在此,我们对来自中国北方的9位鲜卑个体(约公元200年至300年)产生了基因组数据。通过结合文献中现有的基因组,我们建立了一个数据库,涵盖了几乎整个鲜卑时期以及他们之前和之后的样本,使我们能够将鲜卑置于时间背景中进行研究。
我们的研究决定性地解决了一个长期存在的假设,支持鲜卑起源于黑龙江地区,更具体地说是来自中国东北部大兴安岭山脉周边地区。我们还提供了直接的遗传证据,表明在他们最初向中原地区南迁的过程中,鲜卑仅从他们遇到的当地人群中接受了有限的外源基因贡献,但在定居中国北方后,鲜卑不仅从游牧部落转变为定居农业人,而且在遗传上也与当地居民融合。
总之,我们的研究代表了对鲜卑起源的首次基因组探索,确认了鲜卑与古代汉族社群之间深刻的历史联系,并阐明了中国北方的动态人口历史。
引言
中国北方位于定居的粟作农业者和游牧民族的交汇处,不仅是众多古代部落联盟的诞生地,也是两个不同社会之间物质文化、农业和技术交流的熔炉(Di Cosmo 1999)。自铁器时代以来,东欧亚草原上的一系列游牧群体在中国北方及其周边地区建立了政治部落,如匈奴、鲜卑和柔然。其中许多部落南迁至中原,甚至建立了自己的王朝。在这些游牧联盟中,鲜卑或许是最著名的,他们建立了北魏王朝(公元386-534年),这是中国北部边境最长寿和最强大的王朝之一(Holcombe 2013)。
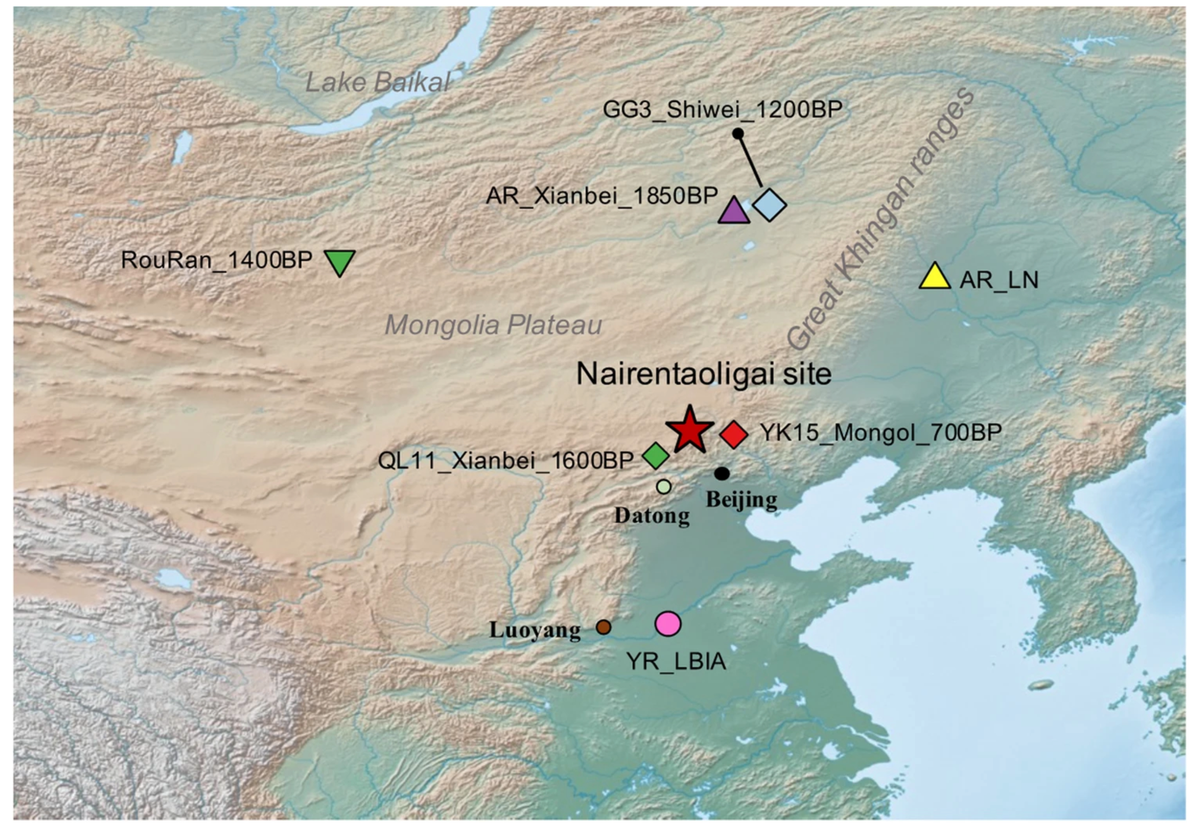
鲜卑首次出现在中国历史记载中是在公元49年的《后汉书》(后汉书 1959),在其全盛时期统治着覆盖整个蒙古高原和中国北方的广大领土。作为一个无文字的社会,关于鲜卑的大多数信息仅来自外部文献,特别是中国史学家的记载;因此,鲜卑的起源仍然存在争议。历史记载表明,鲜卑是古代游牧部落东胡的后裔,后者在公元前206年左右被匈奴击败,随后北迁至"大鲜卑山",即现在被认为是黑龙江地区大兴安岭(后汉书 1959;Holcombe 2013;魏书 1975)。因此,有假说认为广阔的大兴安岭地区是鲜卑的原始家园(图1)。
此外,其他学者还提出了山戎假说和东夷假说,分别将鲜卑的起源追溯到中国东部和北部(Li 1993;Ma 2006)。人类学证据表明,鲜卑可能由多个族群组成,这表明其人口结构较为复杂(Chen等 2003;Chen 2002;Zeng 2009)。由于缺乏文字记录,人们对鲜卑使用的语言知之甚少,现有证据主要来自有限的部落名称(如宇文、慕容和吐谷浑)(魏书 1975),这些证据不足以确定鲜卑语言的分类,尽管大多数学者认为它可能与原始蒙古语有关(Janhunen 2006)。

图1. NRG遗址和已发表的相关遗址的地理位置
在匈奴帝国崩溃后,鲜卑迅速崛起,开始向西和向南迁徙,最终统治了整个蒙古草原和中国北方的广大地区。作为游牧联盟,鲜卑统一了多个部落,包括匈奴的残余势力、贝加尔湖地区的丁零人以及中国北方的乌桓人。一个显著的例子是,在公元91年,鲜卑吸收了匈奴人曾经控制的土地,超过10万匈奴战士接受了鲜卑的身份认同(《后汉书》1959;Holcombe 2013)。这一点得到了考古发现和人类学证据的支持(Zhang 2022;Zhu 1989)。
同时,鲜卑的南迁是其历史的重要组成部分。根据历史记载,鲜卑从其初始家园"大鲜卑山"迁移到呼伦贝尔草原和内蒙古,很快向南迁移并最终定居在中原地区。虽然目前发现的许多考古遗址为鲜卑南迁提供了重要信息,但由于缺乏鲜卑人群的直接基因组数据,这些迁徙事件的详细模式和潜在的人口混合事件仍不清楚。
由鲜卑部落联盟中占主导地位的拓跋氏建立的北魏王朝,在隋唐统一中国之前的北方王朝中具有重要影响力(Holcombe 2013)。尽管军事实力强大,北魏统治者还是实施了一系列政策来缓解与定居汉族人口之间的民族紧张关系。这些政策包括采用汉族姓名、推广汉语,以及鼓励鲜卑人与汉族精英家庭通婚(Holcombe 2013)。这些变化加速了鲜卑与古代中国人之间的互动,但与这种文化转型相关的人口变迁尚未得到充分理解。
尽管鲜卑在中国历史上具有深远意义,但我们对他们的了解仍然相对有限,特别是关于他们的起源以及在汉化过程中复杂的人口迁移事件是否伴随着基因混合。近期考古基因学研究的进展为我们提供了大量来自蒙古草原和贝加尔湖地区的基因组数据,以及中国北方不同时期的系列样本,这为追溯鲜卑的根源和重建鲜卑与邻近游牧部落及定居农业人口之间的基因混合提供了宝贵机会(Damgaard等2018;Jeong等2020;Ning等2020)。
在本研究中,我们从内蒙古奈仁陶力盖(NRG)遗址早期鲜卑时期的21个个体中成功提取了9个个体的古代基因组序列。通过将这些新获得的基因组序列与此前发表的鲜卑基因组,以及来自周边地区早期和晚期的基因组数据相结合(J. Li等2018;Ning等2020)(图1),我们建立了一个独特而全面的数据库。这个数据库使我们能够深入了解鲜卑的遗传起源,并在复杂的鲜卑人口迁移背景下揭示人群基因混合的情况。
材料和方法
伦理声明
本研究使用的牙齿样本来自内蒙古自治区文物考古研究所和乌兰察布博物馆。我们获得了该遗址发掘负责人的许可。
古DNA提取和测序
我们按照既定协议从这些遗骸的牙齿和骨骼样本中提取了古DNA(Peltzer等2016;Yang等1998)。使用NEBNext@ UltraII TMDNA Library Prep Kit for Illumina制备双链文库。简而言之,(1)我们将7 μl End Prep反应缓冲液和3 μl End Prep酶混合物加入50 μl DNA提取物中,在20°C孵育40分钟,65°C孵育30分钟;(2)将2.5 μl接头混合物、1 μl连接增强剂和30 μl连接酶主混合物与第一步的DNA结合,在20°C孵育20分钟。随后加入3 μl USER Mix到反应混合物中,在37°C孵育15分钟;(3)然后使用QIAGEN的MinElute PCR纯化试剂盒纯化混合物,收集18 μl DNA用于最终PCR反应;(4)最后,将30 μl Q5 PCR主混合物、1 μl通用PCR引物和1 μl索引引物加入DNA中,按以下循环进行扩增:98°C 30秒,15次98°C 10秒和65°C 75秒,随后在65°C延伸5分钟。所有文库随后在Illumina HiSeq X10平台上进行测序。
数据处理和基因分型
测序后,使用EAGER软件包处理数据(Peltzer等2016)。具体来说,我们首先使用AdapterRemoval去除Illumina接头并合并配对序列(Schubert等2016)。然后,使用BWA的"aln"和"samse"将单个合并的reads比对到人类参考基因组hs37d5(H. Li和Durbin 2009)。对于具有多个文库的个体,我们使用SAMtools的"merge"合并二进制BAM文件。使用Dedup软件去除PCR重复(Peltzer等2016),并使用MapDamage计算DNA reads的错误掺入率(Jonsson等2013)。随后,使用bamUtils中的trimBam功能修剪每个read的5'和3'端的损坏碱基,以最小化死后DNA损伤(Jun等2015)。最后,我们使用SAMtools mpileup和pileupCaller软件通过随机抽样策略从已发表的"1240 K" SNPs中调用拟二倍体基因型(Mathieson等2015)。
分子性别估计和古DNA真实性
通过评估X和Y染色体覆盖度进行NRG个体的分子性别鉴定。X和Y染色体覆盖度大致相等的个体被归类为男性,而Y染色体覆盖度有限的个体被归类为女性。为验证本研究产生的序列的真实性,我们采用了三种方法。首先,我们观察到所有序列数据的3'端G到A的频率增加。此外,使用Schmutzi评估线粒体DNA污染率(Renaud等2015)。最后,我们使用ANGSD软件比较所有男性个体单拷贝X染色体上多态位点之间的错配率来估计核污染(Korneliussen等2014)。
单亲标记、亲缘关系估计和ROH分析
使用Geneious检索每个个体的线粒体一致序列。使用HaploGrep2进一步确定单倍群(Weissensteiner等2016),使用Yleaf包分配Y染色体单倍群(Ralf等2018),该包使用YFull(v10.01)作为单倍群的基础树结构。使用READ软件估计NRG个体之间的亲缘关系(Monroy Kuhn等2018)。使用默认参数的python包hapROH检测我们古代个体的纯合子连续区域(ROH)块(Ringbauer等2020)。
群体遗传分析
为了描述NRG群体的遗传特征,我们首先通过将新产生的数据与先前发表的全球现代和古代群体合并来准备数据集(补充表8、9)。使用EIGENSOFT包中的smpartpca进行主成分分析(PCA),参数设置为lsqproject: YES和shrinkmode: YES(Patterson等2006)。使用ADMIXTOOLS中的qp3Pop和qpDstat程序计算f3和f4统计量(Patterson等2012),使用非洲的Mbuti群体作为外群。使用ADMIXTOOLS软件包中的qpAdm软件估计潜在的源群体及其比例(Patterson等2012),使用一组群体作为外群,包括Mbuti等多个群体。
结果
考古背景和古DNA数据
NRG墓地位于中国北方内蒙古锡林郭勒盟西乌珠穆沁旗西南约30公里处(42.5415°N,114.0634°E)(图1)。该遗址南北长260米,东西长300米;墓地地形呈缓坡,海拔1336米。遗址共有17座长方形竖井墓,这是鲜卑族常见的墓葬形式。其中四座为合葬墓(M4、M6、M9和M17),十座为单人墓,三座墓葬未发现骨骼。考古学家发现了各种各样的随葬品,包括陶器、青铜器、骨器、羊骨、铜饰牌、五铢钱、桦树皮、玛瑙和绿松石珠。值得注意的是,在M4中发现的五铢钱可追溯到东汉桓帝统治时期,约在公元132年至168年之间(图1)。对NRG017个体的放射性碳测年分析得出的年代范围为1709-1819 BP(补充表1)。根据对各种考古证据的综合评估,该墓群估计可追溯到东汉末期的二世纪末,将NRG群体置于鲜卑联盟的早期(Dang 2021)。
我们在吉林大学考古学院专门设计用于古DNA的实验室中从总共21个骨骼元素(牙齿=18个和骨骼=3个)中提取DNA(补充表1)。在初步浅层筛选后,将九个DNA保存较好的个体(内源人类DNA > 4%)进行进一步低覆盖度测序(每个个体0.02-0.5×)用于后续基因组分析。所有样本都显示出古DNA特有的DNA损伤模式,并具有较低水平的线粒体和核污染(< 4%)(补充图1,表1)。然后,我们将新产生的基因组与两个先前发布的参考数据集合并,这些数据集包括来自Human Origins(HO)数据集和"1240 k"数据集的古代和现代个体。合并的HO数据集用于执行全球群体结构分析,包括主成分分析(PCA)和ADMIXTURE,主要是因为它包含更多当今全球群体。1240 K数据集用于其余分析(如f-统计和qpAdm)。
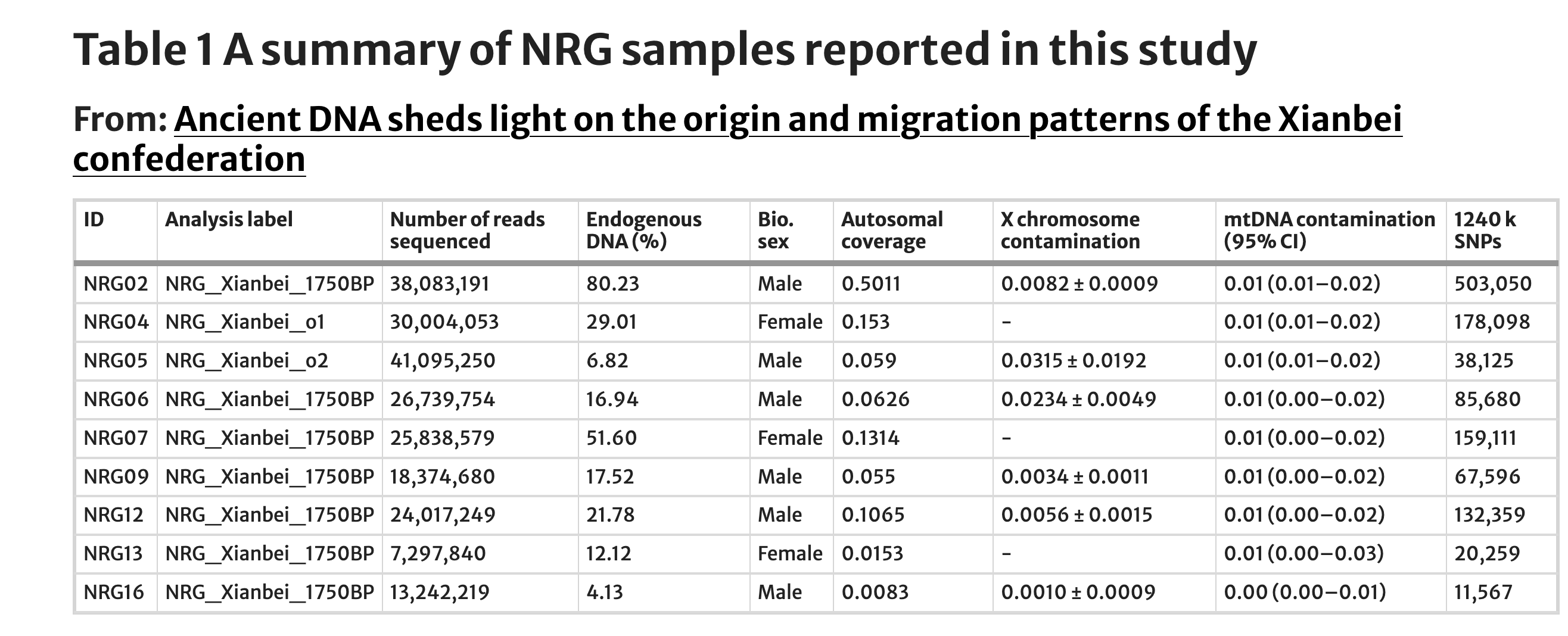
线粒体DNA和Y染色体DNA结果
在21个鲜卑样本中,我们检测到11个主要线粒体DNA单倍群,包括C(C4a1a4和C5d1)、D4(D4、D4a1、D4c2b、D4e1、D4e5a、D4j + 16288和D4m)、M32′56和N9a1。其中,C和D4单倍群在内蒙古蒙古族人群中普遍存在,但M32′56和N9a1在现代当地人群中尚未报道。D4亚型在古代东北亚人群以及阿穆尔河流域的通古斯语系族群(如鄂伦春族和鄂温克族)中普遍存在。C单倍群在东北亚北部被检测到。N9a1单倍群在两个古代个体中被发现,包括西辽河夏家店下层文化晚期新石器时代个体和上游黄河地区铁器时代大草子个体,表明它在古代可能主要分布在中国北方。M32′56单倍群仅在泰国和其他东南亚人群中报道,表明NRG人群可能与东南亚人群有母系遗传关系。
与线粒体DNA单倍群多样性相比,NRG人群的Y染色体单倍群多样性较低。除两个低覆盖度样本外,所有六个NRG鲜卑男性个体都携带C单倍群,其中四个属于亚型C-F1756。C-F1756单倍群在内蒙古当地人群中仅占1.96%,在东亚人群中被检测到。这个单倍群也被三个此前发表的鲜卑个体携带,以及后来被认为是鲜卑后裔的柔然和室韦等游牧民族。综合早期古DNA数据,我们的结果表明C-F1756可能是古代东胡、鲜卑、柔然和室韦部落的主要父系谱系。
亲缘关系估计和纯合片段分析
我们使用READ软件估计了所有NRG个体与此前发表的早期鲜卑个体之间的亲缘关系。READ计算基因组内非重叠1 Mb片段之间的非同源等位基因比率(P0);P0值越低表示共享的染色体片段越多。通过这种方法,我们首先确认九个基因组之间没有完全相同或密切关系的个体。此外,我们重复验证了一对此前发表的一级亲属关系(MGS-M7L和MGS-M7R)鲜卑个体,这与先前的结果一致。结合单亲标记,我们确定他们可能是父子关系。
我们使用hapROH程序进一步评估了鲜卑个体的纯合片段(ROH)。ROH是基因组中缺乏变异的连续区域,这些长的DNA片段的长度可能反映了近亲繁殖的谱系。我们的结果显示,在鲜卑社会中没有近亲繁殖的遗传证据。
NRG鲜卑人群的遗传起源
为了描述NRG鲜卑个体的遗传特征,我们首先对其常染色体基因组数据进行了主成分分析(PCA)。我们将NRG个体以及来自已发表来源的相关古代基因组投射到由现代欧亚人群构建的背景上。我们的结果显示,我们研究中产生的所有9个基因组都落在现代东亚人群观察到的遗传变异范围内。除了两个异常个体外,大多数NRG个体(NRG_Xianbei_1750BP,n = 7)与一个包括最近发表的早期鲜卑样本(AR_Xianbei_1850BP)和阿穆尔河流域新石器时代人群(AR_EN和AR_LN)的群体聚集在一起。这些个体此前被确定为共享一个共同的祖先,统称为古代东北亚(ANA)基因库。他们在地理上分布在大兴安岭周围的阿穆尔河流域地区,表明NRG鲜卑很可能起源于这些地区。
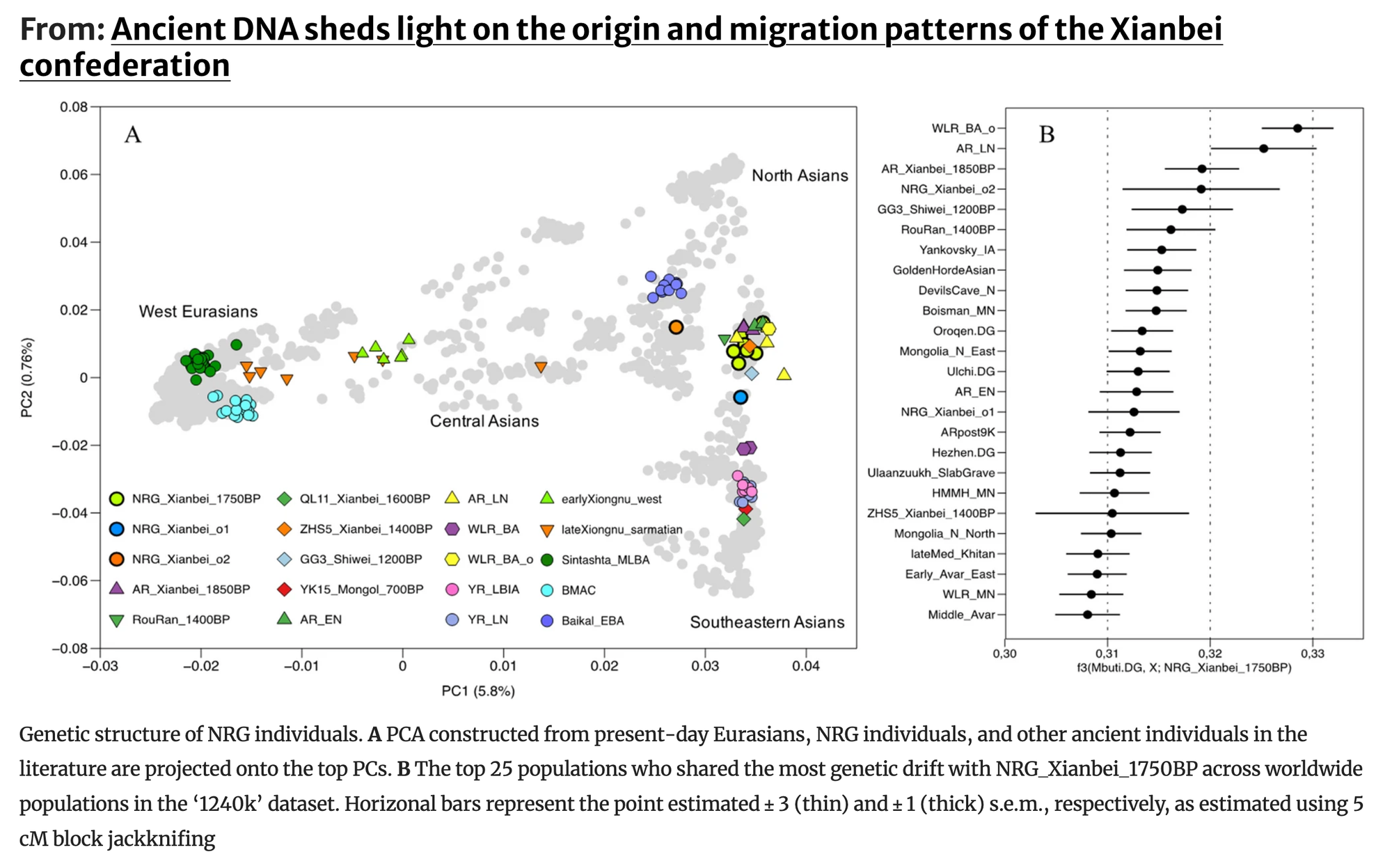
NRG鲜卑群体的遗传亲缘关系和组分分析
我们使用中非姆布蒂人作为外群,通过外群f3统计进一步研究了NRG鲜卑群体与全球人群之间的遗传亲缘关系。我们发现NRG的主要群体(NRG_Xianbei_1750BP)与东北亚古代群体表现出最高程度的等位基因共享。这些群体包括AR_Xianbei_1850BP、AR_LN、GG3_Shiwei_1200BP、Rouran_1400BP和WLR_BA_o,它们都具有相似的古代东北亚(ANA)相关祖先成分。
随后,我们使用qpAdm对NRG_Xianbei_1750BP的潜在祖先成分进行建模。当使用古代游牧人群作为单一来源时,可以通过不同的单向模型充分解释其遗传构成(p值>0.05),如AR_Xianbei_1850BP和AR_LN。我们继续尝试以AR_Xianbei_1850BP为一个来源,其他潜在欧亚人群为另一个来源的双向模型。虽然一些模型显示出适当的p值(p>0.05),但第二个祖先成分的观察比例明显小于标准误差。这一观察结果与主成分分析一致,表明来自中国北方的NRG_Xianbei_1750BP群体与AR_Xianbei_1850BP、AR_LN以及来自大兴安岭周围阿穆尔河流域的其他群体具有很强的遗传亲缘关系。这些发现支持鲜卑可能起源于该地区的大兴安岭假说。
NRG鲜卑人群的遗传混合
除了主要群体外,我们还在NRG群体中发现了两个遗传异常个体。主成分分析结果显示,这两个个体分别与西欧亚人群和阿穆尔河以南的其他东亚个体表现出微弱的遗传亲缘关系。这表明NRG鲜卑人群可能接受了来自周边地区的基因流。
为深入研究这些人口迁移和遗传混合,我们首先使用f4统计形式f4(NRG_Xianbei_o1/NRG_Xianbei_o2,NRG_Xianbei_1750BP;X,Mbuti.DG)。对于NRG_Xianbei_o1,与主成分分析结果一致,我们观察到许多东亚南部人群,如台湾的汉本人群(Taiwan_Hanben)、福建的良渚2号和蒙古的两位汉朝士兵(Han_2200BP),都产生了阳性f4统计值(Z>2)。这些结果表明,与NRG_Xianbei_1750BP相比,NRG_Xianbei_o1具有额外的东亚南部人群相关祖先成分。
对于NRG_Xianbei_o2,与其在主成分分析中的位置相一致,f4统计估计显示NRG_Xianbei_o2与古代西欧亚和中亚人群(如Globular_Amphor、Lisakovskiy_MLBA_Alakul和Rookpund_B)之间存在更多的等位基因共享。这些发现表明,与主群体相比,来自西欧亚或邻近欧亚草原地区的额外基因流对鲜卑人群的遗传构成有所贡献,特别是在NRG_Xianbei_o2的情况下。
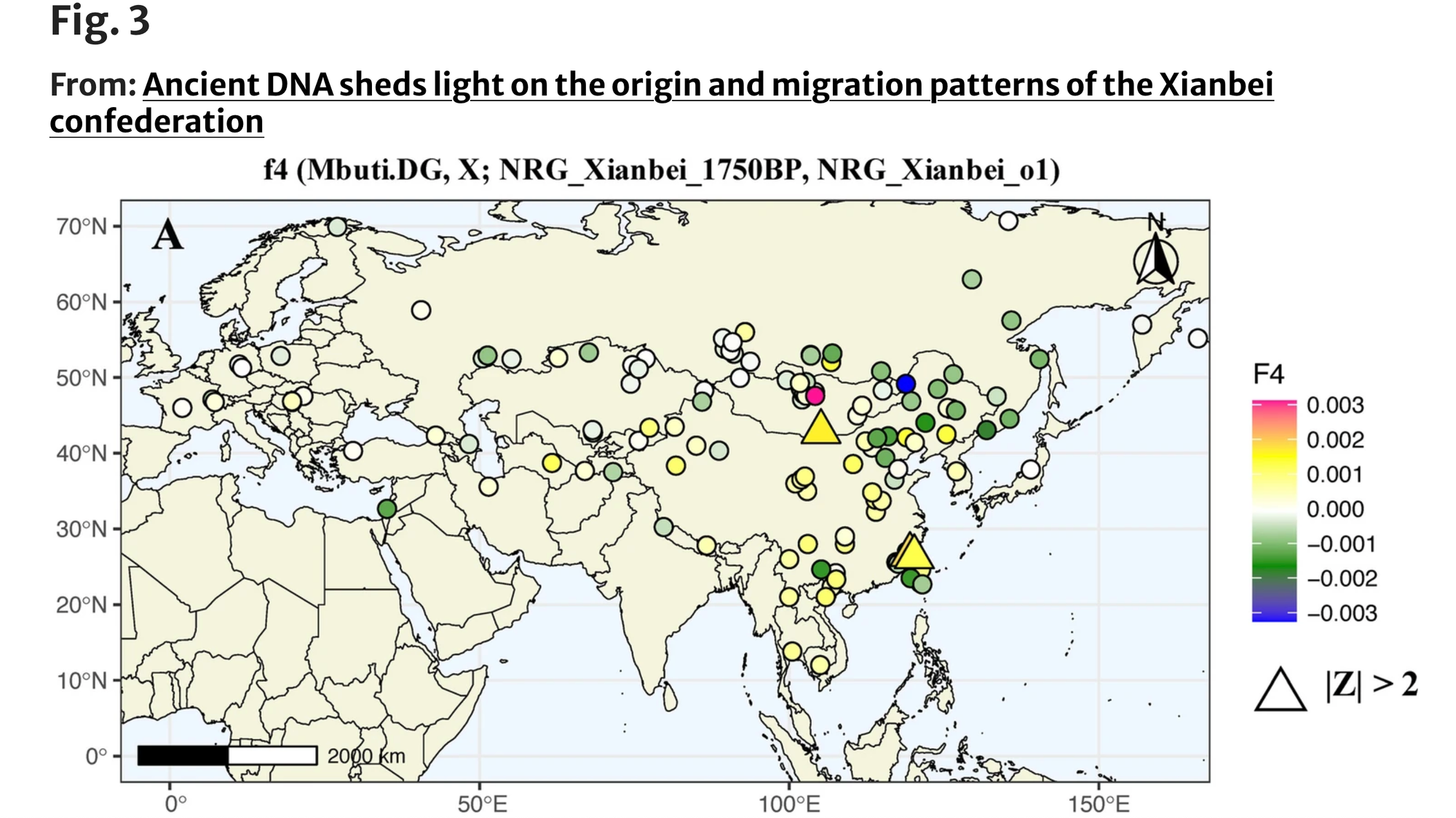
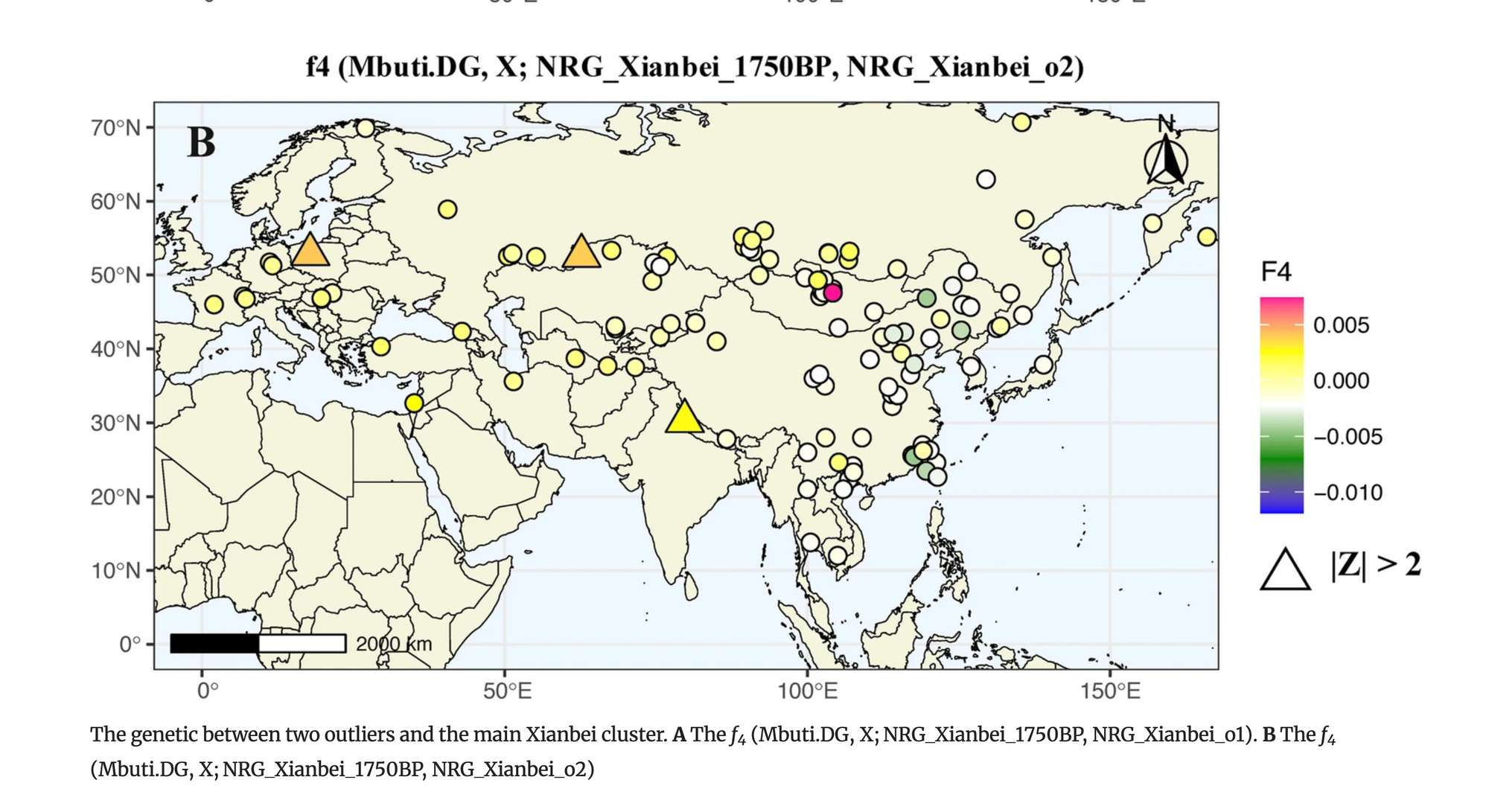
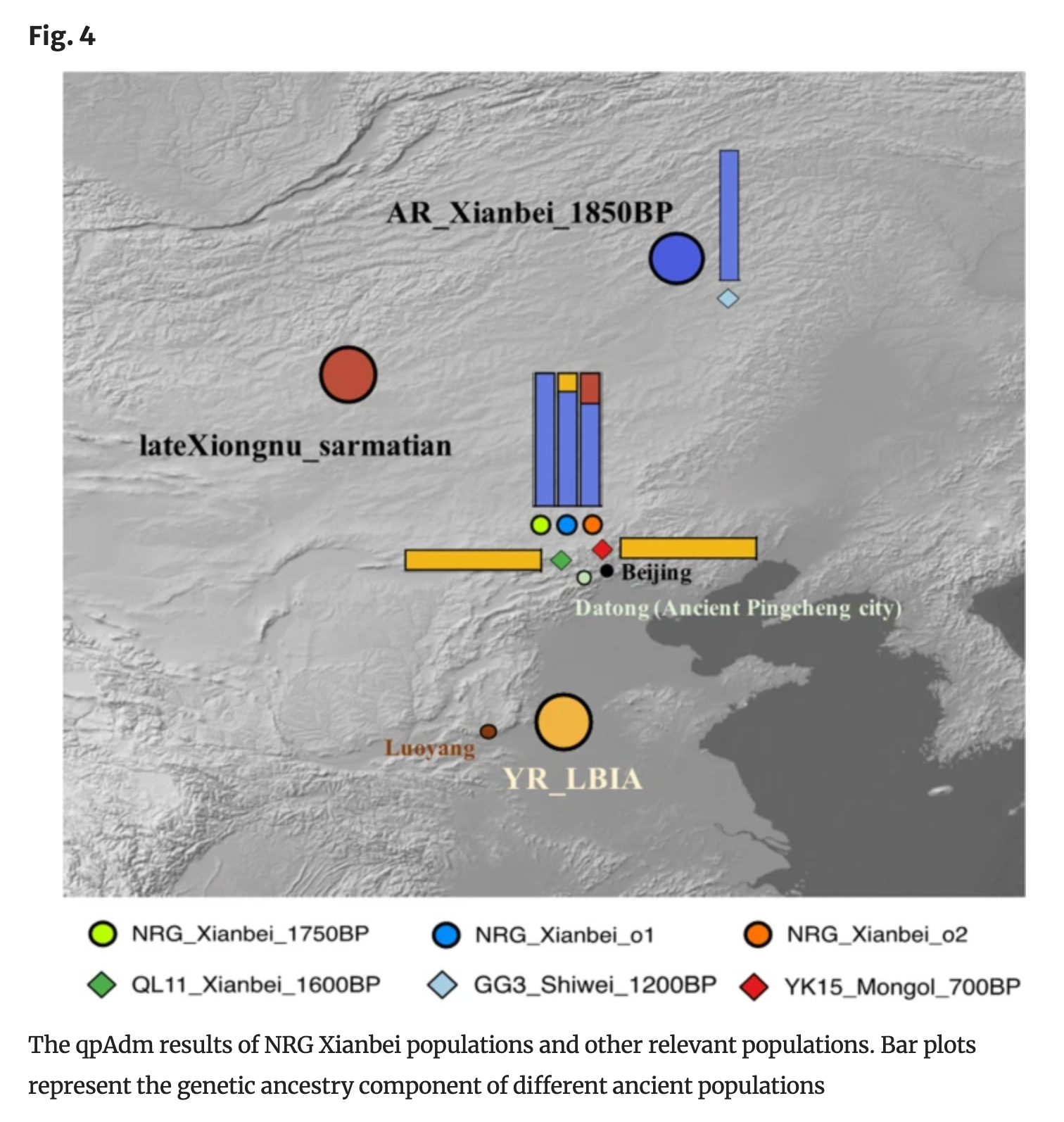
为了更深入地了解这两个遗传异常个体潜在的祖先来源和混合比例,我们使用qpAdm方法对其进行建模。我们选择NRG_Xianbei_1750BP作为一个代表性来源,另外选择了22个可能相关的欧亚古代人群作为另一个来源。总体而言,qpAdm结果表明,这两个异常个体可以被描述为主要与鲜卑相关的祖先成分,以及与周边人群的微弱遗传混合。具体来说,NRG_Xianbei_o2可以通过双向混合模型很好地拟合,其中包括NRG_Xianbei_1750BP(77.4~86.1%)和来自蒙古草原和中亚的多个人群,包括晚期匈奴-萨尔马特人(18.3±6.7%)、早期西部匈奴(22.6±9%)、辛塔什塔中晚期青铜时代人群(13.9±5.3%)和巴克特里亚-马吉安娜考古复合体(15.9±5.6%)(图4,补充表6A)。与NRG_Xianbei_o2类似,NRG_Xianbei_o1的祖先成分也主要来自NRG_Xianbei_1750BP(86.1%),其余部分则更好地由黄河流域晚期青铜时代-铁器时代人群(YR_LBIA)(13.9%)来表征(图4,补充表6B),这与f4统计结果相一致。
总之,综合分析一致表明,在NRG鲜卑的遗传异常个体中存在来自中亚和东亚南部的显著但微弱的祖先贡献。这些发现表明,在NRG时期,鲜卑确实与周边人群发生了遗传混合。重要的是,这些结果与考古发现和历史记录相吻合,进一步支持了该地区在那个时期存在复杂的人口互动和迁徙的观点。
鲜卑南迁后与中原地区日益增加的遗传混合
鲜卑从其最初的家园大兴安岭地区南迁后,其中最具影响力的拓跋部在公元386年在内蒙古中部建立了北魏王朝。最终,他们定都平城,即今天的山西省大同市(《魏书》1975)。在五世纪北魏统治平城期间,鲜卑与古代汉族之间发生了重要的文化交流(Holcombe 2013)。在此期间,鲜卑在经济实践、文化特征和丧葬习俗方面都经历了重大变化。这些互动和转变标志着鲜卑人历史上的一个重要时期,以及他们与更广泛的中华文明的融合(Li 2011; Liu 2020; Ni 2020)。
在五世纪中期北魏王朝崩溃后,中国北方仍然是见证各种历史记载的游牧势力兴衰的地区,如室韦和蒙古帝国。在此期间,定居农业人口与游牧部落之间进行了商品、技术和思想的交流,这对人口流动产生了深远的影响。为了理解这些重大变化及其遗传影响,我们重新分析了此前发表的平城时期的鲜卑基因组(QL11_Xianbei_1600BP)以及中国北方的后鲜卑游牧个体(GG3_Shiwei_1200BP和YK15_Mongol_700BP)(Li等,2020)。
我们首先观察到,单个QL11_Xianbei_1600BP个体显示出与早期鲜卑人群(如NRG个体和AR_Xianbei_1850BP)明显不同的遗传特征,而是落入由中原地区古代黄河农民(如YR_LBIA和YR_LN)形成的群集中,这表明来自中原地区古代农业社群的祖先基因流入在增加。这种与中原地区古代汉族农业社群的强烈亲缘关系也反映在外群f3统计中,其中QL11_Xianbei_1600BP与现代苗族、畲族以及来自中国的古代庙子沟_MN、YR_LBIA和YR_LN等南方东亚人群显示出最强的遗传亲缘关系。
相比之下,来自东蒙古的单个室韦个体(GG3_Shiwei_1200BP)显示出截然不同的遗传特征,在PCA图中与NRG_Xianbei_1750BP和AR_Xianbei_1850BP聚集在一起,表明具有强烈的东北亚亲缘关系。f4统计显示这个个体与主要NRG群体形成一个分支,进一步的qpAdm分析也表明GG3_Shiwei_1200BP可以成功地用NRG_Xianbei_1750BP作为单一来源进行建模。
更晚期的YK15_Mongol_700BP个体与蒙古文化相关,显示出与先前的室韦个体的明显不连续性,在PCA中落入由QL11_Xianbei_1600BP和来自中原的YR_LBIA形成的群集中,并且可以在qpAdm分析中使用YR_LBIA作为代理来源进行单向混合建模,这表明自晚期鲜卑时期(以QL11_Xianbei_1600BP为代表)以来,来自中原地区的遗传影响持续存在且强烈。
讨论
鲜卑在东欧亚草原的重要性已得到历史记录的充分证实,特别是作为首个在中国北方建立帝国的游牧联盟,鲜卑在其全盛时期几乎控制了整个中国北方地区和广大的蒙古高原。然而,鲜卑的遗传起源及其与周边人群的关系仍然存在谜团。在本研究中,我们展示了来自内蒙古早期鲜卑时期的九个鲜卑个体的新基因组数据,并将这些数据与此前发表的来自东欧亚的古代基因组相结合,组建了一个包括鲜卑前后时期人群的数据集。通过我们的新研究,我们追踪了鲜卑的遗传起源,描述了他们的遗传结构,更重要的是理解了鲜卑南迁期间与当地人群的遗传交流,以及从游牧部落转变为定居农业者过程中文化同化与遗传交流之间的相互作用。
首先,鲜卑人的起源一直是研究者们不确定的话题。一些研究者认为他们起源于"鲜卑山",即现在被认定为阿穆尔河地区的大兴安岭(《后汉书》1959;《魏书》1975)。而其他人则假设来自中国东部或北部的人群是鲜卑的祖先(Li 1993; Ma 2006)。此前的基因组研究显示,在中国东北呼伦贝尔地区的1850 BP早期鲜卑个体与阿穆尔河地区的新石器时代猎人-采集者共享同质的遗传特征(Ning等,2020)。我们的研究也表明,鲜卑人群与AR_Xianbei_1850BP以及阿穆尔地区大兴安岭周围新石器时代早期人群显示出最近的遗传联系。这些观察结果也与他们高度同质的父系谱系相符。因此,我们的遗传结果更好地支持了此前提出的大兴安岭假说,而非其他假说。
其次,正如中国史学家所证实的,鲜卑多次南迁,最终定居在接近汉族社会的地方。在这个漫长的迁徙过程中,鲜卑吸收了来自欧亚草原和定居汉族的文化影响。他们还联合了周边地区的各个人群,最终形成了一个强大的游牧联盟。然而,由于缺乏来自古代基因组数据的直接证据,鲜卑与其邻居之间的遗传互动仍然未知。通过结合与鲜卑相关的时间序列数据,我们证明在他们的南迁过程中,以NRG为例,尽管主要NRG群体表现出与早期鲜卑个体相似的遗传特征,但来自邻近地区的遗传影响开始显现。NRG鲜卑中两个接受了来自周边地区少量基因流的遗传异常者,可能是鲜卑与南方农业人群以及蒙古高原游牧民之间互动的结果。此外,我们观察到,与鲜卑个体线粒体DNA单倍群的高度多样性相比,他们的父系谱系相对统一。我们推测,鲜卑男性可能在迁徙过程中与不同的母系人群通婚。
考古证据表明,早在公元一世纪,鲜卑就与周边地区进行了文化和经济交流。例如,在呼伦贝尔的鲜卑墓地中经常出土具有中原特征的漆器、铜镜和陶器(Wei 2004)。此外,鲜卑鹿形饰件的图案可能受到匈奴动物艺术传统的启发,而在NRG发现的汉朝五铢钱表明鲜卑与汉族之间存在广泛的文化互动(Dang 2021; Zhang 2022)。然而,尽管受到汉族的深刻文化影响,我们只在早期鲜卑个体中发现了很小比例的遗传混合。这表明在鲜卑历史的早期阶段,来自周围文化的影响主要是思想的传播。
最后,与早期鲜卑人群相比,1600 BP的鲜卑个体(QL11_Xianbei_1600BP)在遗传上属于黄河流域基因库,该基因库由晚期新石器时代龙山以及河南省青铜时代到铁器时代的个体形成,这揭示了特别是在鲜卑定居中国北方后,来自南方农业人群的直接迁移。这种来自南方的迁移也支持了历史记录中记载的南方文明与北方游牧鲜卑之间广泛的文化联系。公元386年,鲜卑最强大的部族之一拓跋部建立了北魏王朝,定都平城(今山西省大同市)。鲜卑皇帝自称"天子",这是中国传统王朝最高统治者的至高称号,并引入了各种中国式的政治制度,如均田制、中国式官僚制度以及法律和财政制度(Liu 2020; Wei 1975)。
公元494年,孝文帝将大多数贵族、官员和居民从平城迁至新都洛阳(今山西省大同市),并命令拓跋贵族采用汉姓、说汉语、穿汉服、与汉族贵族通婚,并遵循汉族习俗和礼仪(Liu 2020; Wei 1975)。这与鲜卑的稳定同位素分析结果相一致,早期鲜卑以动物蛋白为主食,只有少量粟米消费。但在后期,可能由于受到汉族经济和文化的影响,粟米农业快速发展,成为鲜卑最重要的经济活动之一(G. Zhang等,2015)。我们的研究结果表明,随着文化习俗和饮食习惯的改变,鲜卑也开始接受来自南方农业文明的移民。
结论
我们提供了令人信服的证据,支持鲜卑人起源于阿穆尔地区的大兴安岭。这一证据与此前提出的"山戎"或"东夷"假设起源相矛盾。此外,我们的研究表明,在早期南迁过程中,鲜卑与外部人群的遗传互动有限。然而,当他们在中国北方建立定居点后,我们检测到了来自中原汉族人群的显著遗传影响。特别是在定居中国北方之后,鲜卑的遗传构成越来越接近古代汉族。这一观察结果表明,鲜卑经历了与汉族人群的重要文化和遗传混合。我们还观察到父系遗传的Y染色体多样性低而母系遗传的线粒体DNA多样性高,这可能表明存在父系居住的婚姻模式。然而,必须承认我们目前的分析是基于有限数量的鲜卑基因组。获取更多的鲜卑人群数据将显著有助于我们理解鲜卑社会。
NRG人群的Y染色体单倍群多样性低于线粒体DNA单倍群多样性。除了两个低覆盖度样本外,所有六位NRG鲜卑男性个体都携带C单倍群,其中四位属于C-F1756亚支系(补充表 2)。C-F1756单倍群在内蒙古当地人群中仅占1.96%,主要在东亚人群中被检测到(补充图 2)(https://www.23mofang.com/gene-club/detail/1768930ac9e)(L. H. Wei等,2017),但在三位已发表的鲜卑个体中被发现,以及在后来被认为是鲜卑后裔的古代游牧民族中也有发现,如柔然和室韦(J. Li等,2020;J. Li等,2018;L. H. Wei等,2017)。另外两个个体由于Y染色体SNP覆盖有限,仅被归类为C*(xM217)和C-F1699*(xM48)单倍群(补充表 2)。结合早期的古DNA数据,我们的研究结果表明,C-F1756单倍群可能是古代东胡、鲜卑、柔然和室韦部落的主要父系谱系。然而,需要强调的是,由于本研究中男性个体数量有限,且与鲜卑人群相关的已发表Y染色体数据稀少,对鲜卑人群父系遗传结构的进一步研究将从额外的古DNA数据中获益良多。
 陕公网安备61011302002223号
陕公网安备61011302002223号