古DNA揭示中国新石器时代一个双氏族母系社区
对墓地古DNA的研究为我们了解早期人类社会提供了宝贵的见解,并且强有力地表明了父居制[1](https://www.nature.com/articles/s41586-025-09103-x#ref-CR1),[2](https://www.nature.com/articles/s41586-025-09103-x#ref-CR2),[3](https://www.nature.com/articles/s41586-025-09103-x#ref-CR3),[4](https://www.nature.com/articles/s41586-025-09103-x#ref-CR4),[5

摘要
对墓地古DNA的研究为我们了解早期人类社会提供了宝贵的见解,并且强有力地表明了父居制1,2,3,4,5,6,7,8,9,10。在本研究中,我们分析了中国东部付家考古遗址两处独立墓地中60个个体的古DNA,同时结合考古背景和多种稳定同位素数据,这些墓地年代在公元前2750年至2500年之间。我们的发现表明新石器时代存在一个早期的母系社区,其特点是高度内婚并在沿海地区从事粟作农业。两处墓地之间个体通婚的证据,以及严格按照母系氏族组织的初次埋葬和再次埋葬的存在,凸显了付家遗址强烈的社会凝聚力和认同感。贝叶斯模型的碳14测年表明,这两处墓地使用了大约250年,意味着母系血统稳定延续了至少10代人。这项研究为人类学和考古学中的持续争论11做出了贡献,不仅提示了早期人类历史中母系社会的存在,还揭示了一对围绕两个母系氏族组织的新石器时代墓地,进一步加深了我们通过亲属制度对人类社会早期演化的理解。
引言
早期人类社会是通过父系还是母系血统组织的,这是一个关键但存在争议的问题。在过去的二十年里,考古学家、语言学家和遗传学家为解答这个问题做出了诸多努力,但尚未达成共识12,13,14,15,16,17,18。文化演化的系统发生研究并不支持关于人类社会演化的刚性、单向假说,而是显示出更具动态性和灵活性的亲属制度,这些制度经常发生变化,往往是对环境和经济变化的响应,其中游牧业和农业的传播通常有利于父居制的形成13,19,20。
尽管母系社会组织在现今特别是在中国已经罕见,但它在过去可能更为普遍。近期在遗传分析和语言家族共同祖先的贝叶斯建模方面的进展,为人类社会结构提供了新的见解13,14,15,16,17,18。然而,迄今为止几乎所有研究新石器时代和青铜时代的全基因组研究都一致支持父居制和父系制2,3,4,5,6,7,8,9,10,母系制在远古或史前时期的存在和分布仍然基本未知。到目前为止,基因组分析确认的唯一古代母系社会是北美查科峡谷的精英王朝,年代在公元800年至1300年之间(参考文献21)。在德国南部公元前616年至200年间的一些凯尔特精英中也有可能存在母系血统的迹象22,以及在铁器时代的不列颠,杜罗特里格斯人的墓葬揭示了主导性的母系血统和男性迁入现象23(图1a)。

a,全球人口亲属组织的空间分布,改编自D-PLACE61,62和文献。古代人口的亲属结构用不同的颜色和符号表示,区分了不同地区和时期的母系和父系等各种类型。世界地图使用R语言包maps(v3.4.2)创建。b,付家考古遗址的地理位置与6000年前海侵的关系。标注了与付家同期的周边考古遗址,包括焦家(JJ)、大汶口(DWK)、董家营(DJY)、梁王城(LWC)、西公桥(XGQ)和北前(BQ),以提供背景参照。底图来自World Terrain Base域地图数据集(https://www.arcgis.com/home/item.html?id=c61ad8ab017d49e1a82f580ee1298931),使用ArcGIS v10.2创建。数据来源:Esri、USGS、NOAA。c,付家墓地出土的彩陶(左下),以及南墓地(FJ_S21;左中)和北墓地(FJ_N07;左上)。右侧面板展示了付家考古遗址的布局和特征,突出显示了居住区、陶窑和两处墓地。比例尺,50米。
在本研究中,我们使用优化的古DNA方法获得了全基因组数据,以重建中国东部付家考古遗址公元前2750年至2500年间埋葬的古代个体之间的精确遗传关系。我们的研究结果提供了付家遗址双氏族母系社区的全面基因组证据,超越了仅对线粒体DNA(mtDNA)的高变区进行测序的数据24。我们还采用了跨学科方法,包括稳定同位素分析、骨骼学分类、古环境分析和碳14年代的贝叶斯建模,以深入了解他们的生存方式、当地环境、社会复杂性和墓地使用持续时间。付家案例不仅证实了新石器时代母系社会的存在,填补了古代母系制度证据的空白,还提供了其环境-社会背景,扩展了我们对早期人类社会母系亲属组织多样性和复杂性的理解。
付家遗址的背景和取样
付家遗址(北纬37°2′20.19″,东经118°23′39.63″)位于中国东部山东省,泰沂山脉以北,渤海南岸附近(图1b)。通过考古调查和发掘,付家被确认为大汶口文化晚期遗址(图1c和补充说明1)。遗址面积约37公顷25,26,27(补充表1和补充图1),分别于1985年、1995年和2021年进行了三次发掘(参考文献26)。付家遗址位于靠近海边的低地中的一处高地上,居住区集中在中心地带,陶窑主要分布在西侧的一处小高地上。在遗址的北部和东南部分别发掘出两处独立的墓地,分别标记为Fujia_N和Fujia_S(图1c)。该遗址已发掘出500多座墓葬,现有碳14测年数据显示年代在公元前2750年至2500年之间(补充表2和补充图3)。对墓地加速器质谱测年数据的贝叶斯建模表明,埋葬活动持续了约250年,跨越至少10代人(补充表3和4,补充图4和5以及补充说明2)。
骨骼学分类为我们理解付家的埋葬习俗提供了洞见。在Fujia_S的295位已识别个体中,58%为男性,42%为女性。在Fujia_S的所有355位埋葬个体中,35%被确认为二次埋葬,涉及多具骨骼的合葬占12%。根据1995年发掘的骨骼年龄估计,各年龄段分布均匀(平均年龄23.4岁),表明付家的埋葬习俗没有年龄偏好(补充说明1)。值得注意的是,Fujia_S中一位35-45岁的男性头骨显示出开颅手术的痕迹28,29。
在本研究中,我们选取了66位头骨完整的个体(Fujia_N 15位,Fujia_S 51位;补充数据1)。经古DNA认证后,由于DNA覆盖度过低或现代人DNA污染程度过高,6位个体被从数据集中移除,最终得到Fujia_N 14位和Fujia_S 46位个体的有效数据。群体遗传分析显示这些个体之间具有遗传同质性(补充图6-8和补充说明3)。我们分析了32个人牙釉质样本的锶同位素比值(87Sr/86Sr),并与研究区域内考古动物遗存和现代野生植物新获得的同位素数据进行了对比。此外,我们从人骨和牙本质胶原蛋白中获得了29个经认证的碳(δ13C,其中δ13C = [(13C/12C)样本/(13C/12C)标准] − 1)和氮(δ15N)同位素值。19位个体的新碳14测年数据已报告(补充数据2)。
付家的母系氏族社区
对母系遗传的线粒体DNA和父系遗传的Y染色体的研究表明,墓地位置与线粒体DNA单倍型之间存在明显的对应关系,这种关系与个体的遗传性别无关(图2a)。所有14位来自Fujia_N的个体(3位男性;11位女性)都携带线粒体DNA单倍群M8a3,辛普森多样性指数(SDI)值为0。相比之下,在Fujia_S,一个不同的线粒体DNA单倍群D5b1b占主导地位,46位个体中有44位(15位男性;29位女性)携带此单倍群(95.65%),SDI值为0.08(图2b,c)。此外,埋葬在同一墓地且共享相同线粒体单倍型的个体也展示出相同的线粒体DNA序列,表明他们具有共同的母系血统(补充数据3)。虽然Fujia_S的其余两位个体S17和S27属于线粒体单倍群M8a3,但他们的线粒体DNA序列与Fujia_N个体相差6个碱基,表明这是一个不同的母系血统(补充数据3)。与观察到的线粒体DNA模式相比,Y染色体分析结果显示出显著的单倍型多样性(图2b,c,扩展数据图1和补充数据1)。具体而言,对13个Y染色体覆盖度充足(>0.2×)的男性样本(Fujia_N 3位;Fujia_S 10位)的详细检查使我们能够确定Y染色体单倍群。这项分析揭示,来自Fujia_S的10位男性个体展示出显著的高谱系多样性,识别出6个独特的Y谱系(SDI = 0.89;图2b,c和补充数据1),而来自Fujia_N的3位男性个体表现出3个不同的Y单倍群(SDI = 1),这3个序列与Fujia_S中识别的序列无法区分。通过整合线粒体DNA和Y染色体分析,我们提供了证据表明付家的大多数个体,无论其性别如何,都是按照其母系血统进行埋葬的。
图2:付家个体的单亲遗传标记和生物学亲缘关系。
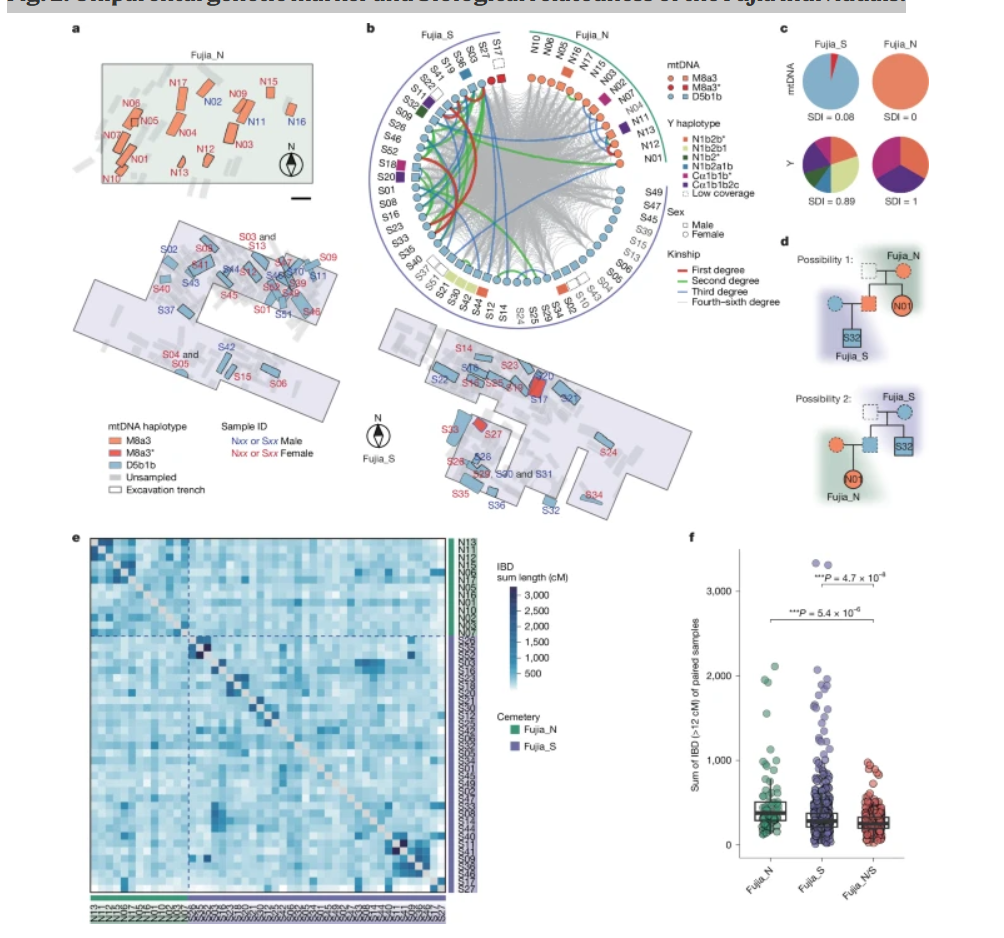
a,付家遗址出土墓葬的放大视图,显示了采样个体的空间分布,根据其线粒体DNA单倍型进行颜色编码,并标注了每个个体的遗传性别。比例尺2米。b,付家个体的遗传关系。最内圈表示每个个体的遗传性别:方形代表男性,圆形代表女性,以及用颜色标示的线粒体单倍型。中间层表示男性个体的Y染色体单倍型。最外层显示每个个体的ID。内圈中的连线说明个体间的遗传亲缘关系,从KIN估算一至三级关系,从ancIBD估算四至六级关系。灰色样本名代表从IBD分析中排除的个体。c,饼图显示线粒体DNA和Y染色体单倍型的组成和频率,突出显示低线粒体DNA多样性和高Y染色体多样性。d,遵循母系血统埋葬的实例,涉及一对二级亲属N01和S32,他们具有不同的线粒体DNA。在这两种情况下,他们的埋葬都与从谱系推断的各自母系血统相对应。e,热图显示具有超过240,000个单核苷酸多态性的个体(n = 44)之间的成对IBD共享,说明Fujia_N和Fujia_S之间存在广泛的亲缘关系。f,墓地间组(Fujia_N,n = 78对;Fujia_S,n = 561对)和墓地内组(Fujia_N/S,n = 442对)的个体对之间的成对IBD共享,揭示了同一墓地内更紧密的生物学亲缘关系。箱形图显示中位数(中心线)、第一和第三四分位数(下和上铰链),以及从铰链延伸至1.5倍四分位距的须线。*P值 < 0.001,来自双侧t检验。
为了了解这些个体之间的亲缘关系,我们对本研究中的60位个体进行了全面的遗传亲缘关系分析。我们使用READ来识别一级遗传关系(扩展数据图2a、补充图9、补充数据4和补充说明4),使用KIN来检测至三级亲属并区分兄弟姐妹和父母-子女关系(图2b和扩展数据图2b),并使用ancIBD来发现至六级的生物学亲缘关系(图2b和扩展数据图2c)。我们的分析发现N01和S32是叔侄关系(图2d和补充说明4),还有两对三级关系,N01和S51,以及N04和S32(图2b)。这些发现进一步证实,这种紧密的遗传亲缘关系并未打破母系埋葬习俗。此外,来自Fujia_S和Fujia_N的所有符合质量标准的个体都表现出较高密度的同源DNA片段(IBD)共享(>12 cM),累积长度从28到975 cM不等(中位数为255 cM;图2e、扩展数据图3c和补充图10)。超过96.8%(1,081对中的1,047对)的个体对共享超过100 cM的IBD片段(补充数据4),表明存在跨越四至六级关系的家族联系。尽管两个分隔墓地的个体之间存在广泛的遗传亲缘关系,且存在二次埋葬现象(补充数据1,表明最初的埋葬不一定在付家遗址),但基于母系的埋葬习俗仍然保持严格一致。
墓地内部比较显示出比墓地之间比较更长的IBD长度和更多的二级亲属(图2b,f),表明同一母系内的个体之间具有比两个不同母系之间更紧密的生物学亲缘关系。考虑到付家墓地中大量的人类遗骸和延长的使用时间(超过200年),以及母系的统一性和父系的高度多样性,可以推断付家墓地是为母系氏族(或母系世系群体)而设,而不是母系大家庭或家族群体。对付家个体的骨骼年龄估计也显示,青少年和成年男性个体都专门埋葬在其本族母系的墓地中(补充数据1)。这种做法符合母系社会的普遍规范,与父系和父居制系统不同,在父系和父居制系统中,女性个体通常与其男性配偶一起埋葬。此外,每个墓地内个体的埋葬位置与遗传距离无关。即使是同一墓地中的一级亲属也不一定被安置在附近(扩展数据图3a-c、补充图11和补充数据5)。这些结果表明,埋葬位置主要由母系氏族归属决定,而不是性别、年龄、亲属关系或遗传瓶颈等因素。因此,我们认为Fujia_S和Fujia_N之间不同的母系血统反映了基于母系氏族身份的有意识的组织结构。
付家社会中的高度地方内婚
对47位个体(34位来自Fujia_S和13位来自Fujia_N)的进一步研究揭示了显著的高频率纯合子片段(ROH;图3a)。具体而言,所有个体都表现出短ROH(<8 cM),总长度在10到127 cM之间,40位(85%)个体的总长度超过40 cM。ROH的总长度主要来自短期和中期ROH(4-20 cM;扩展数据图4a)。这种ROH分布类似于模拟数据中规模有限的人群预期模式(图3b和扩展数据图4b)。我们的结果表明,由于基因库有限导致付家社区内频繁通婚,造成了高度背景亲缘关系,这也导致有效人口规模(Ne)估计大大减少。与其他古代东亚人群相比,付家人群明显较小的Ne(Fujia_S和Fujia_N都在200-400左右;图3c)似乎不是由地理隔离造成的,因为在山东地区发现了大量同期的考古遗址(图1b),而主要是由于社区内通婚习俗造成的基因库减少。对于具有长ROH片段(>20 cM)的基因组的个体,只有47位中的4位显示超过100 cM,表明近亲通婚(如堂表兄妹或更近关系)的频率较低(补充数据6)。除这四位个体外,约47%(n = 22)至少表现出一个超过20 cM的ROH片段,这可能是由于具有相对较远亲缘关系(如二或三堂表兄妹)的个体之间频繁通婚造成的。结合短ROH的分布(补充图12),近亲通婚并非首选的婚姻模式,而是小型内婚社区中不可避免的现象。
图3:付家个体的ROH和有效人口规模估计。
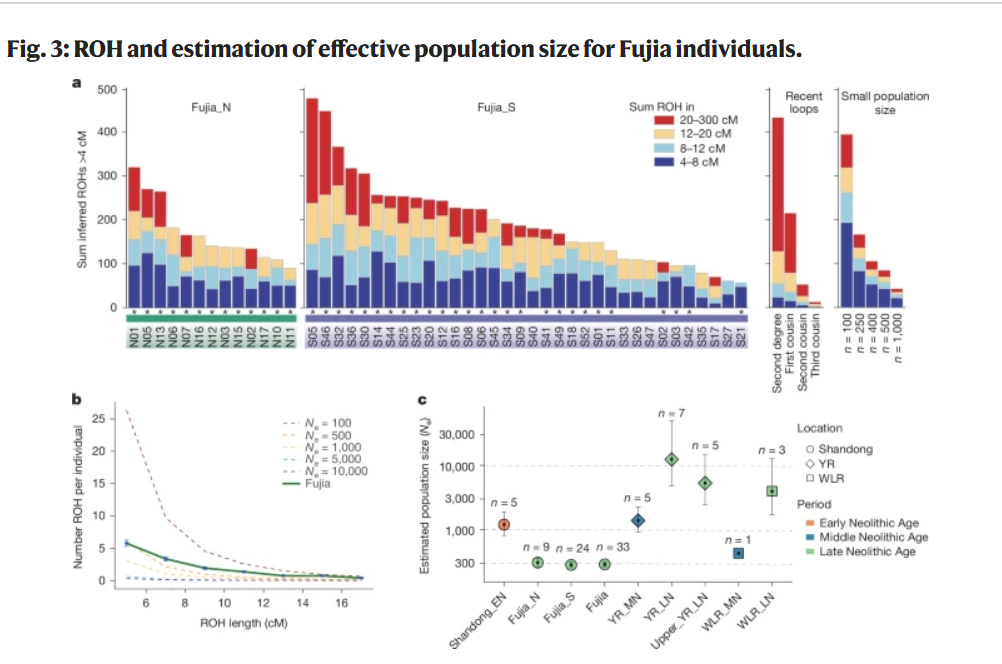
纳入与1240k基因分型位点重叠且具有超过200,000个单核苷酸多态性的个体。a,展示了付家北区和付家南区个体推断的纯合子片段(ROH)。为二级亲属到一至三代表亲之间的近亲个体,以及来自不同规模小群体的个体提供了预期值。付家的ROH长度分布与小群体规模的预期相符。星号表示短ROH片段(<8 cM)累积长度超过40 cM的个体。b,付家个体(n = 33)在各长度类别中的ROH片段平均数量。基于存在长度超过20 cM且总长度超过50 cM的ROH片段,排除了14位高度近亲个体。垂直误差线表示每个长度区间(2 cM)的±1标准误。虚线展示了不同有效人口规模(Ne)的预期ROH分布。c,付家和其他古代东亚人群的估计有效人口规模,表明付家具有较小的人口规模。黑点表示最大似然估计,垂直误差线表示95%置信区间。样本量(每个遗址分析的个体数量)标注在每个对应点上方。YR,黄河;WLR,西辽河;EN,早期新石器时代;MN,中期新石器时代;LN,晚期新石器时代。
付家生存策略和流动性
来自付家的52位个体的δ13C值范围在-9.3‰至-6.6‰之间,平均值为-7.9 ± 0.5‰(本研究29个,之前研究36个样本23个),表明其以C4植物为主的饮食结构。主要的食物来源可能包括粟(Setaria italica)、黍(Panicum miliaceum)和以谷物饲养的猪,考古植物学证据表明更多依赖于以粟为基础的食物。
人类的δ15N值范围在8.1‰至10.5‰之间(n = 52;平均9.3 ± 0.5‰),与付家猪(7.0 ± 0.6‰)相比偏差约2.3‰,表明其饮食中包含以C4植物饲养的动物蛋白。在付家或其他中期或晚期大汶口遗址中,男性和女性个体之间的δ13C或δ15N值没有显著差异,表明不同性别之间具有统一的饮食习惯。
值得注意的是,付家人类的δ15N值显著高于其他大汶口遗址。这种升高可能反映了晚期大汶口时期该遗址沿海环境中盐碱土壤条件的影响,和/或海洋和淡水资源的摄入,这一点从附近遗址(如吴村)发现的海洋软体动物以及付家和其他大汶口遗址发现的淡水贝类得到证实。这些发现共同表明,付家居民从事以粟作农业和畜牧业为主,并可能获取海洋和/或淡水资源。
图4:付家个体的稳定同位素分析。
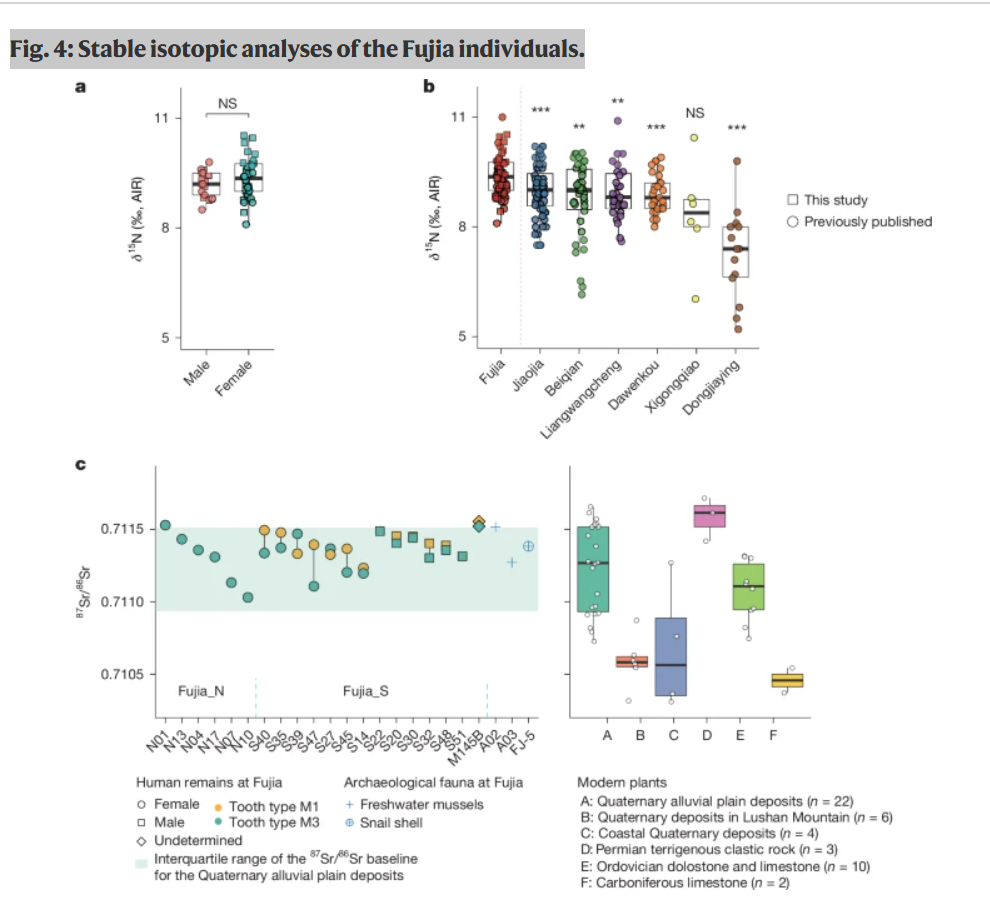
a,付家男性(n = 17)和女性(n = 34)个体骨骼和牙本质胶原中的δ15N值,通过双侧t检验显示没有显著差异,且未进行多重比较调整。AIR为大气空气标准。b,付家与大汶口文化遗址之间的δ15N值比较。双侧t检验比较了付家(n = 51)与六个同期遗址:焦家(n = 60,P = 5.9 × 10−4)、北前(n = 38,P = 0.0012)、梁王城(n = 27,P = 0.0076)、大汶口(n = 29,P = 2.9 × 10−4)、西公桥(n = 7,P = 0.13)和东郊营(n = 14,P = 1.9 × 10−5;补充数据2)。***P值 < 0.001;**P值 < 0.01;NS,无显著差异;未进行多重比较调整。c,付家个体(n = 20)牙釉质中87Sr/86Sr比值与来自不同岩性单元的野生植物(n = 53)和考古动物样本(n = 3)的值进行比较。对于所有面板中的箱线图,中心线表示中位数值;须线表示1.5倍四分位距;点表示观察值。
对20位个体(包括12位具有成对臼齿的个体)进行的锶同位素分析显示,87Sr/86Sr比值范围非常窄,在0.711030至0.711554之间(图4c和补充数据2)。这一范围完全位于付家遗址所在的第四纪冲积平原沉积物的基线范围内(由当地野生植物确定),并且与该地区其他地质单元的范围明显不同(图4c),表明所有付家个体可能都居住在遗址周围的第四纪冲积平原沉积物区域内。尽管一些个体的87Sr/86Sr值与附近山区的奥陶纪白云岩和石灰岩重叠,但付家人群中氧同位素的最小变化(1标准差 < 1‰,n = 23)进一步表明这些个体并非来自高海拔地区,而是共享同一水源(详见补充说明5)。
在第四纪冲积平原沉积物的更局部范围内,所有个体的87Sr/86Sr值都落在考古遗址10公里半径范围内采集的植物范围内(≈0.7107-0.71153),并且与付家出土的蜗牛壳和淡水贝类的87Sr/86Sr值非常接近(图4c),这表明付家人群可能具有有限的居住流动性。这些发现进一步支持付家人群可能在地理上受到限制的观点,与中国其他同期或后期新石器时代群体相比,没有明显的远距离通婚影响。均匀的氧同位素特征和在大汶口人群中最窄的饮食生态位范围进一步表明付家社区具有本地化的活动范围和相对统一的生计实践。基因分析证实了这些发现,表明存在内婚习俗和可能的凝聚社区(补充说明5)。虽然男性个体通常表现出略窄的87Sr/86Sr范围,但使用Mann-Whitney检验未发现统计学上的显著差异(P值 = 0.149)。
讨论
一个多世纪前,社会学家如Johann Jakob Bachofen、Lewis Henry Morgan和Friedrich Engels提出,古代社会普遍是母系的,随着私有财产的出现才演变为父系结构。自二十世纪以来,人类历史早期母系阶段的假说一直受到社会人类学家的激烈争论。一些人认为母系阶段的想法是基于希腊神话和推断,其他人则指出在其他灵长类动物研究中缺乏支持母系社会的证据,也缺乏直接的考古证据。现在,我们可以使用更有说服力的科学方法重新审视这个话题,结合新的数据和方法,特别是与古代基因组相关的方法。
我们的结果表明,付家社区可能是围绕母系原则组织的。这一解释得到了遗传特征与墓地边界之间强烈对应关系的支持。埋葬在同一墓地的个体不仅共享相同的线粒体单倍型,而且在大多数情况下,还具有相同的一致性线粒体DNA序列,表明存在密切的母系血统,可能还有共同的母系身份。这两个墓地之间观察到的二至六级亲缘关系连接的高比例(60.9%,644对中的392对)也表明两个墓地长期共存,使用时间超过约250年。这些模式与现代母系社会的模式相似,后者通常由多个母系氏族组成(如走婚制的摩梭人、妻居制的拉祜族和特林吉特人),其中个体终身保持与其出生母系氏族的关系,并相应地进行埋葬。
 陕公网安备61011302002223号
陕公网安备61011302002223号