欧亚大陆东部草原6000年的动态基因史
[https://www.cell.com/cell/fulltext/S0092-8674(20)31321-0?_returnURL=https%3A%2F%2Flinkinghub.elsevier.com%2Fretrieve%2Fpii%2FS0092867420313210%3Fshowall%3Dtrue](https://www.cell.com/cell/fulltext/S0092-8674(20)31321-0?_returnURL=https%3A%2F%2Flinkinghub.elsevier.com%2Fretrieve%2Fpii%2FS009286742031
Mongolia_Khovd_MLBAXiongnuLateEarlyMedieval_o_KHI001.AG-KHI001 (1).txt
科布多的历史景观:从中晚青铜时代到晚期中世纪
蒙古国科布多地区的历史景观是由从中晚青铜时代、匈奴时期直至晚期中世纪时期在该地区涌现的多样化且充满活力的文化所编织而成的历史画卷。这一区域坐落于雄伟的阿尔泰山脉和广阔的草原之间,曾是一系列游牧文化的发源地,这些文化的影响力曾经在整个中亚及其他地区产生深远影响。
中晚青铜时代(约公元前2000-1000年)
在中晚青铜时代,科布多地区以发展复杂社会和丰富的物质文化为特征。这一时期见证了独特考古文化的出现,如鹿石-石冢文化(Deer Stone-Khirigsuur culture),以其雕刻有鹿纹图案的立石和广泛分布的墓葬丘(石冢)而闻名。这些文物表明当时社会拥有复杂的宗教或精神信仰,与自然和土地有着深厚的联系。
这一时期的社区主要以游牧为生,依靠马术和畜牧业。马、牛、山羊、绵羊和骆驼是他们经济和生活方式的核心。培育和管理大型畜群的能力促进了广泛的贸易网络的形成,使这些社会能够参与到跨越欧亚大陆的更广泛互动中。
青铜时代冶金技术的进步导致了各种工具和武器的生产。铜和锡合金青铜的引入使得工具更加坚固耐用,支持了农业和军事活动。
匈奴联盟(约公元前3世纪-公元1世纪)
在公元前3世纪末,包括科布多地区在内的蒙古草原成为匈奴联盟的摇篮,这是最早的原蒙古游牧帝国之一。这一联盟通常被视为中国南方王朝,特别是汉朝的强大对手。匈奴建立了一个复杂的多民族联盟,控制了中亚广大地区。
匈奴的政治结构相当复杂,采用联邦制使各部落在统一单一单于(最高统治者)的同时保持一定程度的自治。这种中央集权的领导对组织军事行动和管理与邻国的外交关系至关重要。
匈奴是熟练的骑手和弓箭手,这些技能是他们军事成功的关键。他们的游牧生活方式,加上复合弓和骑兵战术的使用,使他们成为一支令人生畏的力量。匈奴还与汉朝进行贸易,交换马匹、皮毛和其他商品以换取丝绸和粮食。
考古发现,包括墓葬和文物如陶器、武器和珠宝,提供了关于他们社会结构、仪式以及与其他游牧部落和定居文化联系的见解。
晚期中世纪(约11-14世纪)
到了晚期中世纪,科布多地区继续成为欧亚大陆文化和商业交流的重要走廊。这一时期包括各种突厥和蒙古部落的兴起和影响,每个部落都为草原文化的丰富镶嵌画增添了色彩。
13世纪初由成吉思汗领导的蒙古帝国对科布多地区的影响尤为显著。在蒙古人统治下,该地区被整合进历史上最大的连续帝国之一,通过蒙古和平(Pax Mongolica)促进了不同文化之间前所未有的交流。这一时期见证了贸易的繁荣和技术、思想、宗教的交流,丝绸之路活动增加。
与蒙古大部分地区一样,科布多在这一时期深受引入的佛教习俗影响。这种精神影响反映在考古遗址和历史文本中,展示了传统萨满教习俗和密宗佛教之间的融合。
从蒙古帝国到随后的分裂王国,最终到各个汗国的过渡,权力动态发生了变化,但该地区作为文化交流熔炉的角色一直保持不变。
结论
从青铜时代到晚期中世纪,蒙古科布多地区成为游牧草原社会的重要文化和经济中心。该地区的历史因一系列有影响力的文化而显著,每种文化都为当代蒙古所见证的遗产做出了贡献。其战略位置和民众持久的适应能力凸显了科布多在中亚历史叙事中的重要性。
蒙克海尔汗县:青铜时代考古重要发现的故乡
蒙古国科布多省的蒙克海尔汗县是青铜时代遗址的丰富区域,特别是在考古学中被称为"蒙克海尔汗文化"的独特遗迹。这一文化于2003年由蒙古-俄罗斯联合的"中亚考古"项目研究队在蒙克海尔汗县境内、蒙克海尔汗山脚下首次发现并发掘研究,从而被引入科学研究领域。
**蒙克海尔汗文化:**属于早期青铜时代的这一文化遗址主要有两种基本类型:墓葬和祭祀设施。墓葬通常呈圆形或正方形,其特点是不会从地表突出,而是被平石覆盖。
**出土文物:**蒙克海尔汗文化墓葬中发现的物品包含了关于当时人们生活和习俗的宝贵信息。其中包括:
- **青铜刀:**带木柄、单刃、尖端略微弯曲的短青铜刀。
- **青铜针:**同样带木柄、方形尖端的青铜针。
- **骨勺:**可能用于埋葬仪式或日常使用的骨制勺子。
- **装饰品:**白石珠、项链等装饰品反映了当时人们的审美观念。
这些文物通常放置在人体膝盖和大腿周围。
**分布范围:**蒙克海尔汗文化遗址不仅限于蒙克海尔汗县,目前已确认分布在巴彦-乌列盖、科布多、戈壁-阿尔泰、扎布汗、库苏古尔、布尔干等省份的广大地区。
**科布多省其他青铜时代遗址:**作为科布多省的一部分,蒙克海尔汗县还有机会发现该地区广泛分布的其他青铜时代遗迹,如石冢、方形墓葬、鹿石和岩画等。特别是沿蒙古阿尔泰山脉的地区一直是古代游牧民族文化的摇篮,蒙克海尔汗县构成了这一历史遗产的重要部分。
查找蒙克海尔汗县及其相关蒙克海尔汗文化(Munkhkhairkhan culture)考古报告、研究成果需要几个特定来源,您通常需要在学术机构数据库和数字图书馆中搜索。
这些报告很少有直接"下载"链接,通常以学术论文形式发表。建议您在以下地方搜索:
1. 蒙古科学院考古研究所:
- 网站:http://www.archaeology.mas.ac.mn/
- **出版物:该研究所出版的"Studia Archaeologica"**学术期刊期刊中可能发表了关于蒙克海尔汗文化的文章。这些出版物可在研究所图书馆找到,有时也会以数字形式放在网站上。
- **主要研究人员:**专门搜索首次发现该文化的D.Tseveendorj、A.P.Derevyanko、V.V.Volkov等研究人员的作品会很有效。
2. 学术论文集成数据库:
研究人员在这些网站上发布自己的作品,您可以下载PDF版本。搜索时使用以下关键词:
- "Munkhkhairkhan culture"
- "蒙克海尔汗文化"
- "Bronze Age Mongolia"
- "D.Tseveendorj archaeology"
- Academia.edu:https://www.academia.edu/ - 许多蒙古和外国考古学家在这里发布他们的作品。这是最有可能的来源之一。
- ResearchGate:[链接已删除] - 另一个科学家互相联系、分享作品的平台。
3. 国外大学、研究中心:
由于蒙克海尔汗文化是由蒙古-俄罗斯联合考察队发现的,可以在俄罗斯科学院西伯利亚分院考古与民族学研究所(Институт археологии и этнографии СО РАН)网站上搜索。
- 俄罗斯科学院西伯利亚分院考古与民族学研究所:http://www.archaeology.nsc.ru/ - 可以从该研究所的出版物、报告中找到相关信息。
4. Google Scholar:
- Google Scholar:https://scholar.google.com/ - 提供在所有类型的学术作品中一站式搜索的功能。使用上述关键词搜索会出现相关书籍、论文列表,其中一些可以免费下载。
具体例子:
例如,D.Tseveendorj、V.V.Volkov、E.A.Okladnikova等合著的**"蒙克海尔汗青铜时代文化"**等标题的文章,或蒙古-俄罗斯联合考察队的报告。这些通常发表在学术期刊上,因此您需要找到相应期刊号。
结论:
考古完整报告通常不对公众开放,而是存放在研究所内部档案馆、图书馆中。而基于报告的研究文章、书籍可以在上述Academia.edu、ResearchGate、Google Scholar等平台和科学院考古研究所出版物中搜索,这是最有效的方法。
亮点:
• 对蒙古和贝加尔地区214个古代个体进行的全基因组分析
• 蒙古晚青铜时代存在三个基因上明显不同的乳制品游牧群体
• 匈奴游牧帝国通过不同本地和远距离群体的混合而形成
• 尽管有5,000年的乳制品文化,但未发现对乳糖酶持续性等位基因的选择压力
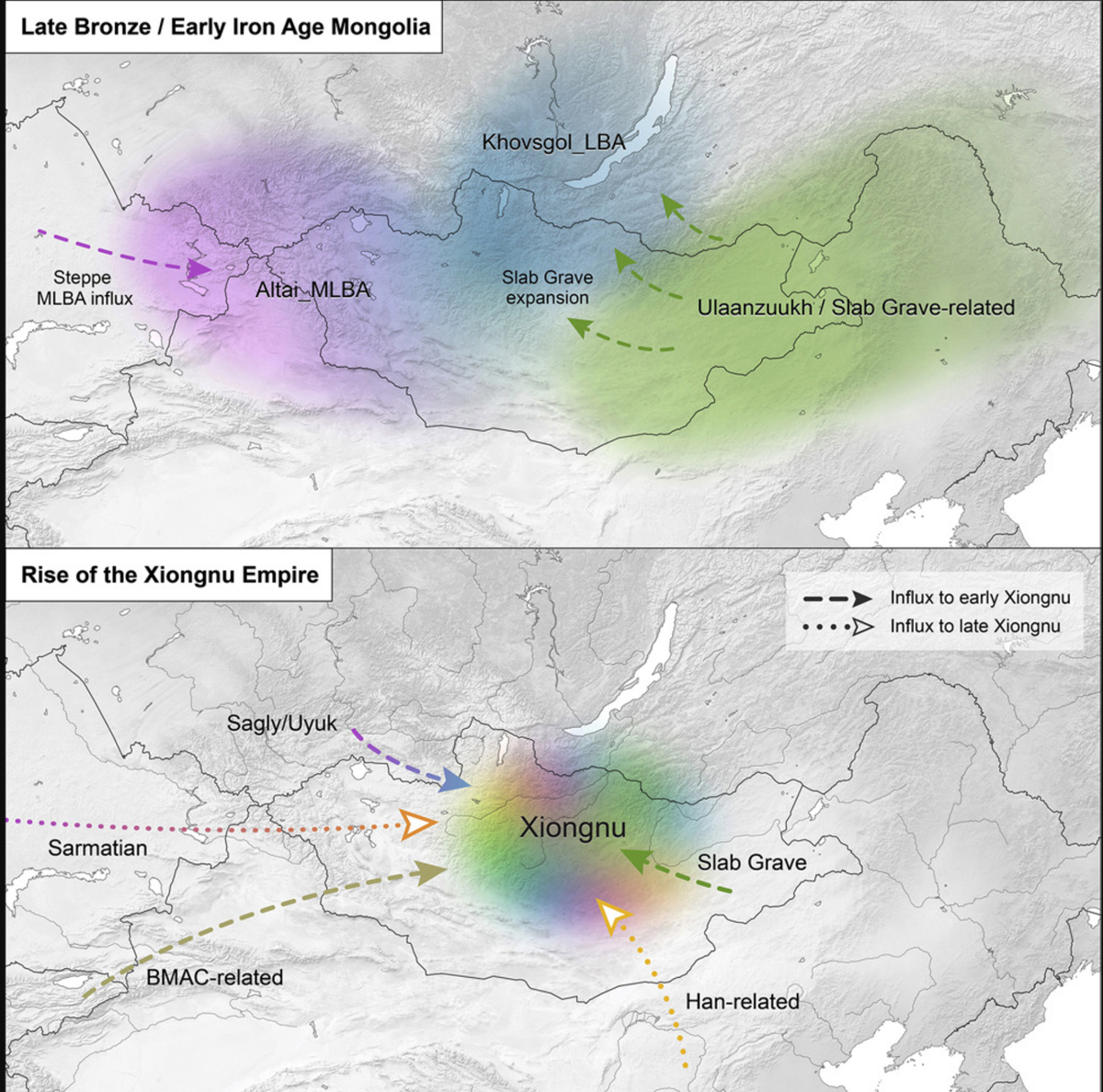
概要:东部欧亚草原是游牧民族历史帝国的故乡,包括匈奴和蒙古人。然而,对该地区的人口历史知之甚少。在此,我们通过分析214个跨越6,000年的古代个体的全基因组数据,揭示了其动态的基因历史。我们确定了大约公元前3000年牧民向蒙古的扩张,到了晚青铜时代,蒙古人口在生物地理上分为三个截然不同的群体,无论血统如何,所有群体都实行乳制品游牧业。匈奴帝国由这些人口与周边地区人口的混合而形成。相比之下,蒙古人表现出更高的东欧亚血统,类似于现今说蒙古语的人口。我们的研究结果阐明了东部草原上基因、社会政治和文化变化之间的复杂相互作用。
关键词
引言
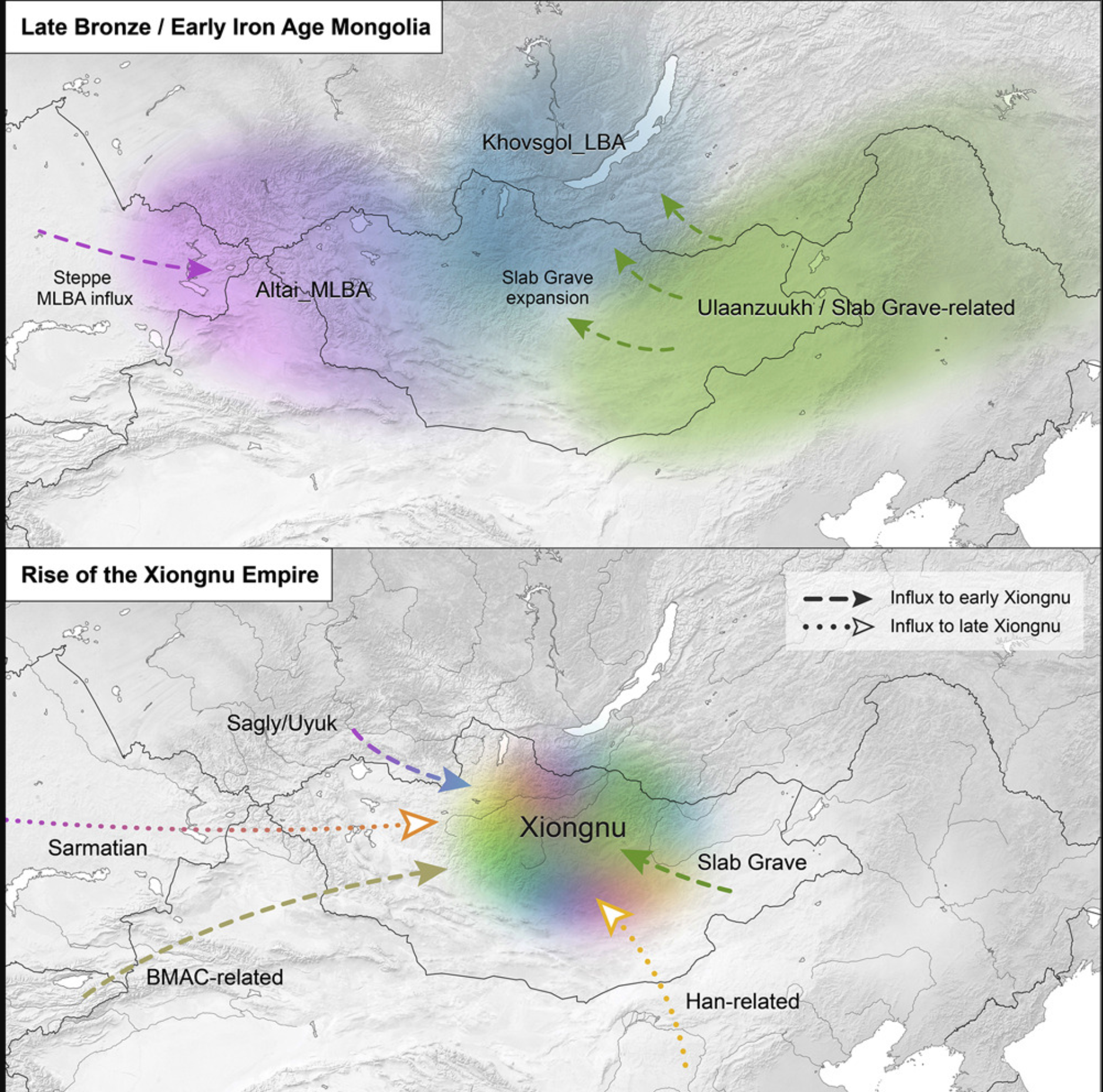
近期的古基因组学研究揭示了欧亚草原上动态的人口历史,西部草原上大陆规模的迁徙事件与青铜时代欧洲、近东和高加索地区的转变相吻合(Allentoft et al., 2015; Damgaard et al., 2018a; 2018b; Haak et al., 2015; Mathieson et al., 2015; Wang et al., 2019)。然而,尽管在理解西部草原的遗传史方面取得了进展,东部草原的史前人口动态仍然知之甚少(Damgaard et al., 2018a; Jeong et al., 2018; Rogers, 2016)。东部草原是一片广阔的草原、森林草原和荒漠草原,绵延超过2,500公里(图1;图S1)。虽然也覆盖了现代中国和俄罗斯的部分地区,但东部草原的大部分落在现今蒙古国的国界内。最近的古基因组学研究表明,东欧亚森林草原带在前青铜时代和早期青铜时代期间在基因上具有结构性,存在一条强烈的西-东混合血统梯度,从哈萨克斯坦中部的博泰延伸到西伯利亚南部的贝加尔湖,再到俄罗斯远东的魔鬼门洞穴(Damgaard et al., 2018a; Jeong et al., 2018; Sikora et al., 2019; Siska et al., 2017)。

图1 古代人口和时期概览
(A) 遗址分布及其相关文化和时期以颜色标示:前青铜时代,紫色;早期青铜时代,红色;中/晚期青铜时代,蓝色;早期铁器时代,粉色;匈奴时期,绿色;早期中世纪,棕色;晚期中世纪,金色(参见研究方法)。有关遗址代码和标签,请参见图S1A和表S1B。
(B) 欧亚大陆插图地图,标示了本研究的区域(方框)以及文中提到的其他古代人口的位置,按时期着色。西部/中部草原的地理范围以浅棕色表示,东部草原以浅绿色表示。
(C) 蒙古主要时期和考古文化的时间线。遗址位置已经进行了抖动处理,以提高重叠遗址的可见性。
图S1 蒙古国考古遗址与地理和生态特征,与图1相关
(A) 本研究分析的蒙古国及周边地区考古遗址。
(B) 蒙古国地区和省份(aimags)。省份以灰色线条和文字标示。地区以黑色虚线和文字标示,遵循(Taylor et al., 2019)的定义。
(C) 蒙古国生态区域。地图使用QGIS软件(v3.6)制作,生态数据来源于(Dorjgotiv, 2004)。
在青铜时代,多阶段引入的牧业生活方式极大地改变了东部草原的生活方式和生存手段(Honeychurch, 2015;Kindstedt and Ser-Od, 2019)。最近一项大规模的古蛋白质组学研究证实,在公元前2500年之前,阿凡纳谢沃文化(约公元前3000年)和切穆尔切克文化(公元前2750-1900年)的个体已经在蒙古地区饮用牛奶(Wilkin et al., 2020a)。虽然上叶尼塞河地区的阿凡纳谢沃群体在基因上与彭蒂克-里海草原的亚姆纳亚文化(约公元前3300-2200年)有联系(Allentoft et al., 2015;Morgunova and Khokhlova, 2013;Narasimhan et al., 2019),但切穆尔切克文化的起源一直存在争议(Kovalev, 2014)。一旦引入,反刍动物的乳制品生产到中/晚期青铜时代(MLBA,这里定义为公元前1900-900年)变得广泛,在西部和北部与鹿石-石冢复合体(DSKC)相关的遗址以及东部与乌兰祖赫文化相关的地区都有实践(Jeong et al., 2018;Wilkin et al., 2020a)。DSKC和乌兰祖赫群体之间的关系尚不清楚,对蒙古其他MLBA埋葬传统,如蒙克海尔汗和拜塔格,知之甚少。到公元前一千年中期,先前的MLBA文化开始衰落,早期铁器时代文化兴起:东部/南部蒙古的板石墓文化(约公元前1000-300年),其墓葬有时包含从DSKC纪念碑中拔出的材料(Fitzhugh, 2009;Honeychurch, 2015;Tsybiktarov, 2003;Volkov, 2002),以及西北部萨彦山脉的萨格里/乌尤克文化(约公元前500-200年)(也被称为萨格里-巴兹文化,或蒙古的昌德曼文化),它与阿尔泰和东哈萨克斯坦的帕济雷克文化(约公元前500-200年)和萨卡文化(约公元前900-200年)有着强烈的文化联系(Savinov, 2002;Tseveendorj, 2007)。
从公元前1千年末开始,东部草原上出现了一系列等级制度明确、中央组织的帝国,特别是匈奴帝国(公元前209年至公元98年)、突厥帝国(公元552-742年)、回鹘帝国(公元744-840年)和契丹帝国(公元916-1125年)。匈奴帝国是草原上第一个这样的政权,其向中国北部、南西伯利亚和深入中亚的大规模扩张对欧亚大陆的人口结构和地缘政治产生了深远影响。蒙古帝国于十三世纪崛起,是这些政权中最后也是最具扩张性的一个,最终控制了从中国到地中海的广阔领土和贸易路线。然而,由于缺乏大规模的基因研究,组成这些国家的人民(包括统治精英和当地平民)的起源和关系仍然不明确。
为了阐明史前时期以来东部草原地区的人口动态,我们生成并分析了来自85个蒙古和3个俄罗斯遗址的214个个体的全基因组数据集,跨越约6,000年的时间(约公元前4600年至公元1400年)(表S1,S2和S3A)。此外,我们还添加了最近发表的北蒙古19个青铜时代个体的基因组数据(Jeong等,2018),以及来自俄罗斯和哈萨克斯坦邻近古代人群的数据集(Damgaard等,2018a;2018b;Narasimhan等,2019;Sikora等,2019;Unterländer等,2017)(表S3B和S3C),我们将这些数据与全球现代参考人群一起分析(表S3C)。我们还生成了30个新的加速器质谱测年数据,补充了74个先前发表的放射性碳测年数据(Jeong等,2018;Taylor等,2019),本研究共有98个直接测年的个体(共104个测年数据)(表S4)。
结果:史前青铜时代的人口结构和牧业的到来
在本研究中,我们分析了来自三个遗址的六个青铜时代前的狩猎-采集者个体,这些遗址可追溯至公元前五千年和四千年:一个来自蒙古东部(SOU001,"eastMongolia_preBA",公元前4686-4495年校准年代),一个来自蒙古中部(ERM003,"centralMongolia_preBA",公元前3781-3639年校准年代),以及四个来自贝加尔东部地区("Fofonovo_EN")。通过将这些基因组与欧亚大陆先前发表的古代和现代数据进行比较(图2;表S3C),我们发现它们与同时期来自贝加尔西部地区("Baikal_EN",公元前5200-4200年)和俄罗斯远东地区("DevilsCave_N",约公元前5700年)的狩猎-采集者关系最为密切,填补了这一基因谱系分布在地理上的空白(图3A)。我们将这一谱系称为"古东北亚"(ANA),以反映其相对于另一种广泛分布的全新世中期基因谱系"古北欧亚"(ANE)的地理分布,后者见于西伯利亚马尔塔(约24500-24100 BP)和阿丰托瓦戈拉(约16900-16500 BP)遗址的更新世狩猎-采集者(Fu等,2016;Raghavan等,2015)以及哈萨克斯坦博泰的饲马者(约公元前3500-3300年)(Damgaard等,2018a)中。在主成分分析(PCA)中(图2),古代ANA个体接近当今东北亚通古斯语系和尼夫赫语人群的聚类,表明他们的基因谱系在今天的远东土著人群中仍然存在(图S3A)。在基因上,eastMongolia_preBA与ANA群体DevilsCave_N无法区分(图3A和4A;图S4A;表S5A),而Fofonovo_EN和稍晚的centralMongolia_preBA都有少量(12%-17%)的ANE相关(类似Botai)祖源,其余(83%-87%)则为ANA祖源(图3A和4A;表S5A)。重新分析贝加尔西部早期新石器时代基托伊文化(Baikal_EN)和早期青铜时代格拉兹科沃文化(Baikal_EBA)的已发表数据(Damgaard等,2018a),我们发现它们具有相似的祖源谱系,并且ANE祖源随时间略有增加(从6.4%增加到20.1%)(图3A)。
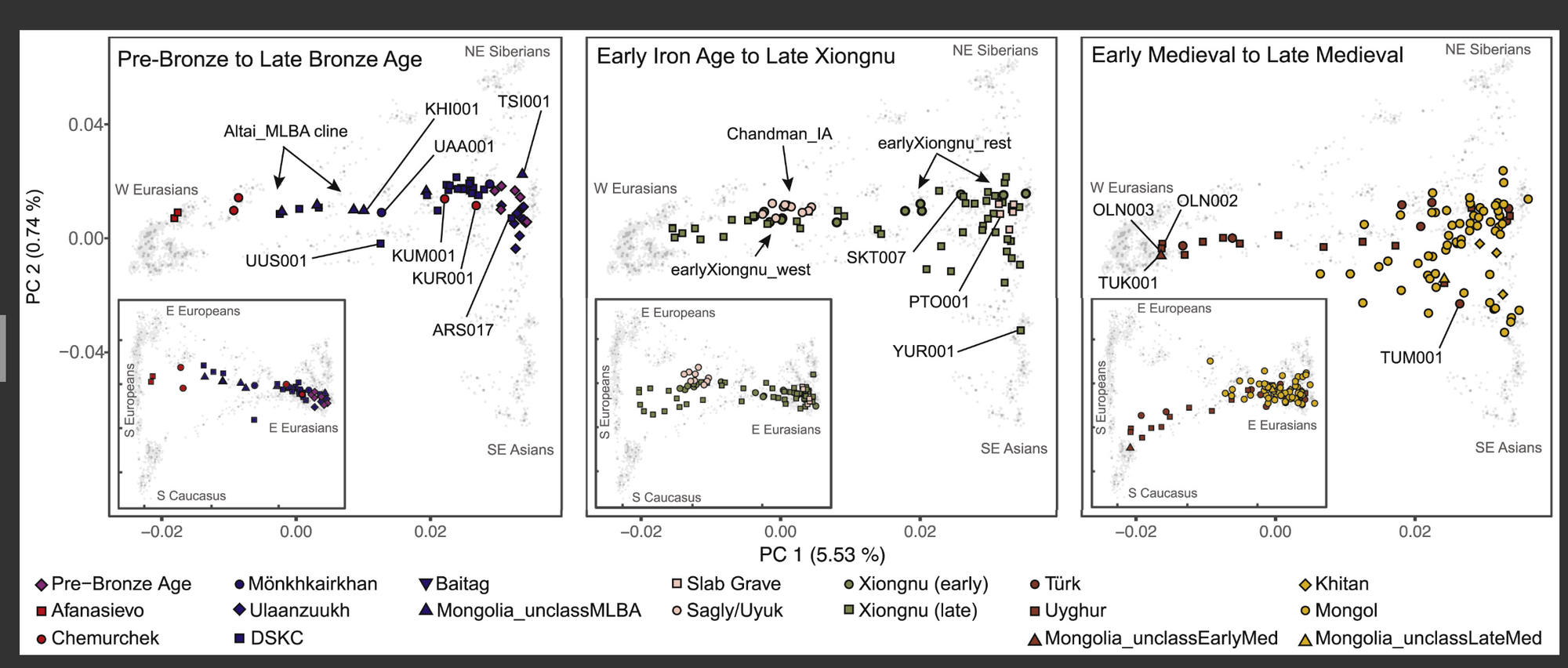
图2 蒙古历史上的遗传结构
古代个体(n = 214)来自三个主要时期,投影到当代欧亚人群(灰色符号)的主成分分析图。主面板显示PC1对PC2;插图显示PC1对PC3。插图中PC1的刻度标记与主面板相对应;PC3解释了0.35%的变异。有关人群、样本和轴标签,请参见图S3B,有关遗址和样本的更多详情,请参见表S1B、S1C和S2A。

图3 通过qpAdm表征的东部草原随时间的基因变化(A–F)主要时期:(A)青铜时代前期至早期青铜时代,(B)中期/晚期青铜时代,(C)早期铁器时代,(D)匈奴时期,(E)早期中世纪,和(F)晚期中世纪。模型化的祖源比例通过按样本大小缩放的饼图表示,祖源人群显示在下方(见研究方法)。每个面板的样本大小范围在右上角标示。对于(B)和(C),Baikal_EBA以浅蓝色建模;在(D–F)中,Khövsgöl_LBA(紫色)和Chandman_IA的Sagly/Uyuk(粉色)被建模为新的源头(图4)。文化群体用粗体文本表示。对于(D–F),个体分别为晚期匈奴、突厥和蒙古,除非另有说明。先前发表的参考人群用白色文字标注;其他所有人群均来自本研究。地图边界以外的人群用箭头表示。埋葬地点已经过抖动处理,以提高重叠个体的可见性。
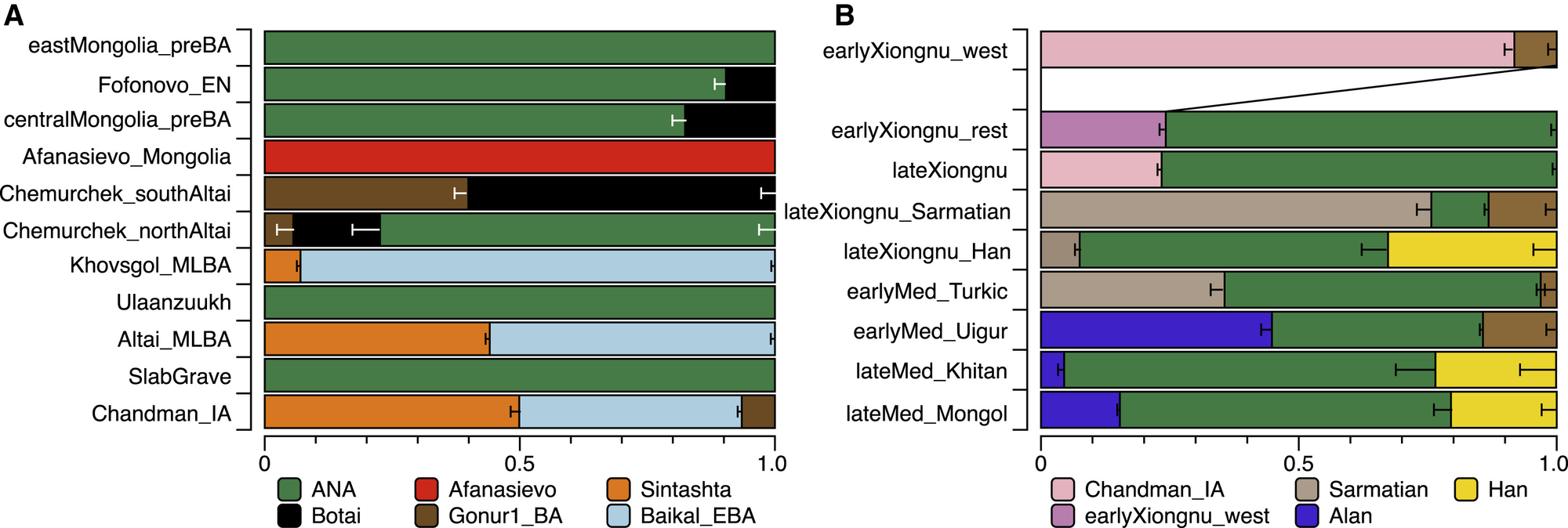
图4 所有新报告的遗传群体按时间顺序的遗传祖源变化基于群体的人口遗传学分析的优良拟合建模结果,包括(A)史前时期和(B)历史时期。每个遗传群体中的个体数量见表S3A。原始祖源比例和标准误差估计见表S5。水平条表示由qpAdm估计的±1标准误差(SE)。
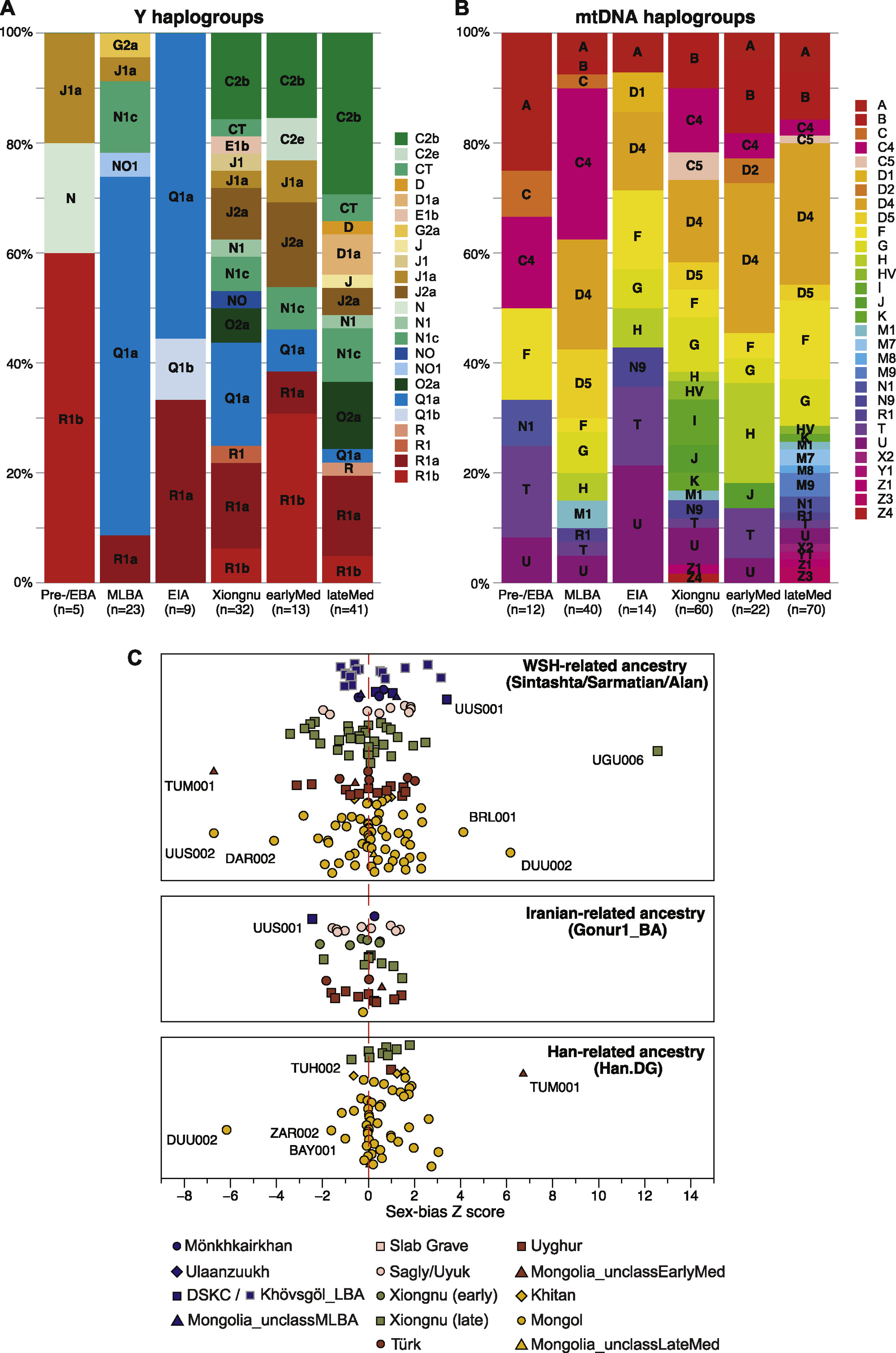
图S2 按群体和性别偏差的单亲单倍体群分配及Z评分,与图5B和STAR方法相关
(A和B) 单亲遗传标记显示的人群结构。(A) 各时期Y染色体单倍体群的分布。(B) 各时期线粒体单倍体群的分布。
(C) 通过评估常染色体和X染色体上WSH-/伊朗-/汉族相关祖源的差异得出的性别偏差Z评分。我们为每个具有这三种祖源之一的遗传混合的古代个体计算Z评分。正值表明常染色体上有更多的WSH-/伊朗-/汉族相关祖源,即男性驱动的混合。

**图S3 当代欧亚人群的主成分分析和蒙古历史上的遗传结构,与图2**相关
(A) 用作图2和图S3B背景的当代欧亚人群主成分分析。这里我们展示了用于计算主成分并在图2中以灰点表示的2,077个欧亚个体的人群标签。图中每个三字母代码代表一个个体。与三字母代码匹配的人群ID列在底部。
(B) 蒙古历史上的遗传结构。来自三个主要时期的古代个体(n = 214)投影到当代欧亚人群(灰色符号)上的主成分分析(PCA)。投影和轴方差对应于图2。人群标签位于属于每个人群的个体的平均坐标上方。

图S4 通过qpAdm表征的东部草原随时间的基因变化,与图3和图4相关
(A) 青铜时代前期至早期青铜时代;(B) 中期/晚期青铜时代;(C) 早期铁器时代;(D) 匈奴时期;(E) 早期中世纪;(F) 晚期中世纪。
模型化的祖源比例通过按样本大小缩放的饼图表示,祖源人群显示在下方。文化群体用粗体文本表示。对于(D–F)面板,个体分别为晚期匈奴、突厥和蒙古,除非另有说明。先前发表的参考人群用白色文字标注;其他所有人群均来自本研究。地图边界以外的人群用箭头表示。埋葬地点已经过抖动处理,以提高重叠个体的可见性。放大以查看个体标签。
我们在此报告的混合模型包含了解释整体晚期中世纪个体所需的所有祖源成分,以便在个体之间进行无偏比较。使用更简单混合模型的个体结果可在表S5J中找到。有关建模详情,请参见第7节。
通常认为蒙古的畜牧业是由西方草原文化(如阿法纳谢沃文化)向东扩张引入的,这一扩张要么通过蒙古西北部的上叶尼塞河和萨彦山脉地区,要么通过西部的阿尔泰山脉(Janz et al., 2017)。虽然目前报道的大多数阿法纳谢沃墓葬位于阿尔泰山脉和上叶尼塞河地区,但中蒙古南部杭爱山脉的早期青铜时代(EBA)遗址沙塔尔楚鲁出土了阿法纳谢沃风格的墓葬,其中有反刍动物乳制品消费的蛋白质组学证据(Wilkin et al., 2020a)和西欧亚线粒体单倍体群(Rogers et al., 2020)。通过分析其中两个个体(蒙古阿法纳谢沃人,公元前3112-2917年校正年代),我们发现他们的基因特征与已发表的叶尼塞河地区阿法纳谢沃个体的基因特征没有明显区别(Allentoft et al., 2015; Narasimhan et al., 2019)(图2;图S5C;表S5B),因此这两个阿法纳谢沃个体证实了早期青铜时代西方草原牧民(WSH)的扩张范围比阿尔泰山向东延伸了1,500公里,深入到中蒙古的腹地(图3A)。
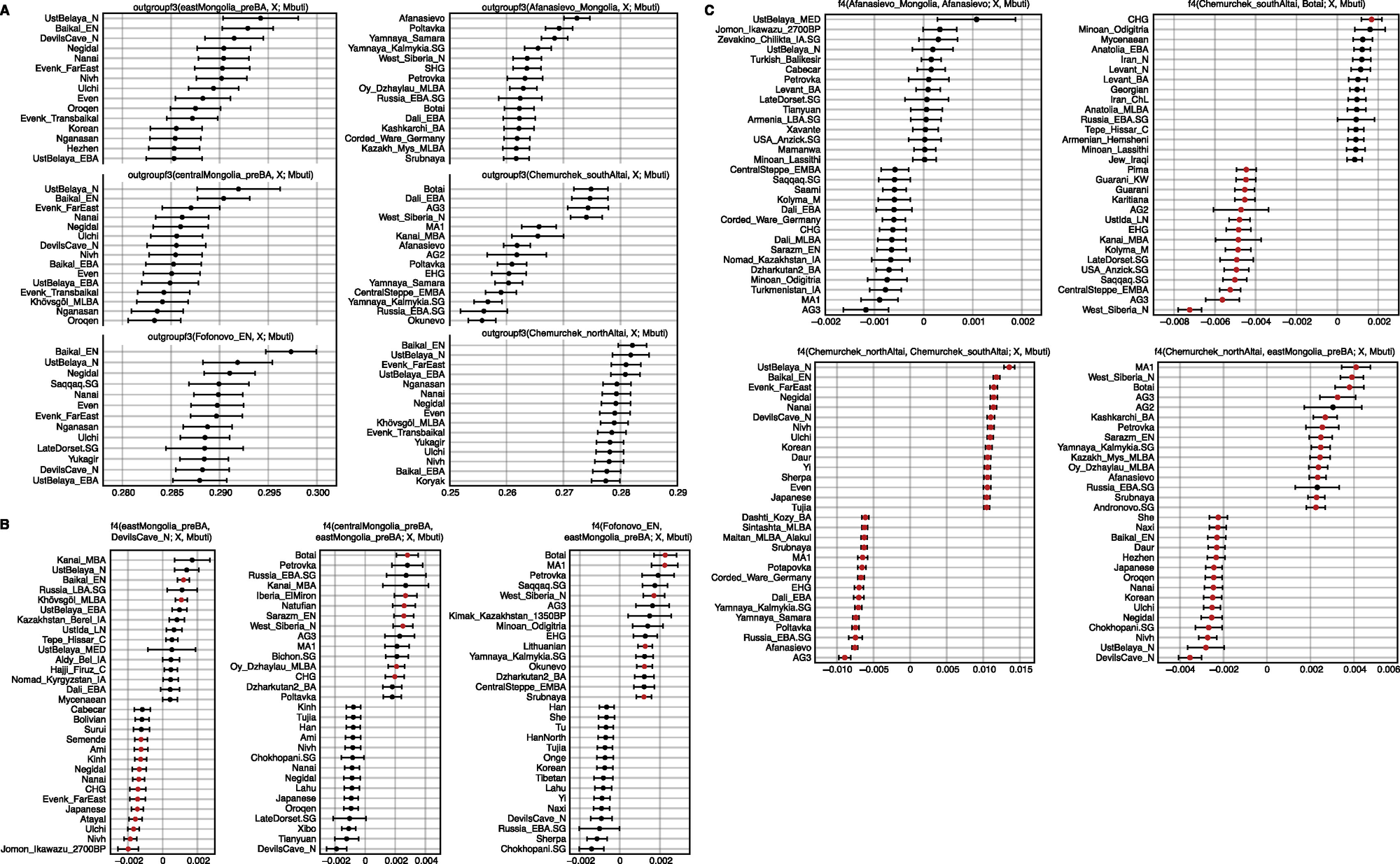
图S5 外群f3-统计和使用f4-统计的分支检验,与图3和4相关
(A) 东部草原青铜时代前至早期青铜时代群体的外群f3-统计。我们展示了六个目标群体的形式为f3(目标群体, 全球范围群体; Mbuti)的前15个外群f3-统计,这些目标群体包括:东蒙古_前青铜时代(eastMongolia_preBA)、中蒙古_前青铜时代(centralMongolia_preBA)、福福诺沃_新石器时代早期(Fofonovo_EN)、蒙古阿法纳谢沃(Afanasievo_Mongolia)、南阿尔泰切穆尔切克(Chemurchek_southAltai)和北阿尔泰切穆尔切克(Chemurchek_northAltai)。水平条表示通过5 cM区块重复抽样计算的±1标准误差(SE)。
(B) 使用f4-统计检验四个ANA人群的分支关系。我们展示了四个与ANA相关的目标群体的形式为f4(ANA1, ANA2; 全球范围群体, Mbuti)的前15个和后15个对称f4-统计,这些目标群体包括:东蒙古_前青铜时代、中蒙古_前青铜时代、福福诺沃_新石器时代早期、魔鬼洞_新石器时代(DevilsCave_N)。水平条表示通过5 cM区块重复抽样计算的±1标准误差(SE)。Z-分数>3的f4-统计以红色突出显示。
(C) 使用f4-统计检验阿法纳谢沃和切穆尔切克的分支关系。我们展示了三个目标群体蒙古阿法纳谢沃、南阿尔泰切穆尔切克和北阿尔泰切穆尔切克的前15个和后15个对称f4-统计,形式分别为f4(蒙古阿法纳谢沃, 阿法纳谢沃; 全球范围群体, Mbuti)、f4(南阿尔泰切穆尔切克, 博泰; 全球范围群体, Mbuti)、f4(北阿尔泰切穆尔切克, 南阿尔泰切穆尔切克; 全球范围群体, Mbuti)和f4(北阿尔泰切穆尔切克, 东蒙古_前青铜时代; 全球范围群体, Mbuti)。水平条表示通过5 cM区块重复抽样计算的±1标准误差(SE)。Z-分数>3的f4-统计以红色突出显示。
随后的早期青铜时代切穆尔切克文化(公元前2750-1900年),是一个反刍动物乳制品社会(Wilkin等,2020a),其墓葬特征包括石板和人形石碑,据推测也与西方草原(WSH)迁徙有关联(Kovalev和Erdenebaatar,2009)。切穆尔切克墓葬遍布阿尔泰地区和中国新疆准噶尔盆地(Jia和Betts,2010;Kovalev,2014;2015)。我们分析了来自南阿尔泰亚格申胡都遗址的两个切穆尔切克个体和来自北阿尔泰昆迪戈壁(KUM001)和库拉伊戈壁2(KUR001)遗址的两个个体。与蒙古阿法纳谢沃人相比,亚格申胡都个体也显示出高度的西方血统,但在主成分分析(PCA)中位置有所偏移(图2),并且与ANE相关的古代个体如阿丰托瓦戈拉3(AG3)、西西伯利亚新石器时代人和博泰人有很强的遗传亲缘关系(图3A;图S5A和S5C)。我们发现亚格申胡都切穆尔切克个体("Chemurchek_southAltai")在遗传上与达利早期青铜时代人("Dali_EBA")相似(图3A),后者是来自哈萨克斯坦东部的同时期个体(Narasimhan等,2019)。亚格申胡都和达利早期青铜时代个体的遗传特征可以用博泰人(60%-78%)和具有古伊朗相关血统的群体(如来自贡努尔台佩的Gonur1_BA,这是巴克特里亚-马尔吉亚纳考古复合体(BMAC)的一个重要早期青铜时代遗址)(22%-40%;图3A;表S5B)的双向混合模型很好地拟合。虽然不能排除阿法纳谢沃相关群体的微小遗传贡献,但所有拟合模型都需要伊朗相关的血统,据DATES模型估计,这种混合发生在12±6代之前(约336±168年;图S6)(Narasimhan等,2019)。然而,由于此建模中使用的所有代理源人群在时间或空间上都与早期青铜时代阿尔泰相当遥远,因此尚无法精确确定切穆尔切克人的近源人群。在北阿尔泰,两个切穆尔切克个体("Chemurchek_northAltai")主要具有ANA衍生的血统(约80%),其余部分类似于南阿尔泰切穆尔切克个体(图3A和4A;表S5B)。因此,我们观察到切穆尔切克个体根据地理位置表现出遗传异质性。
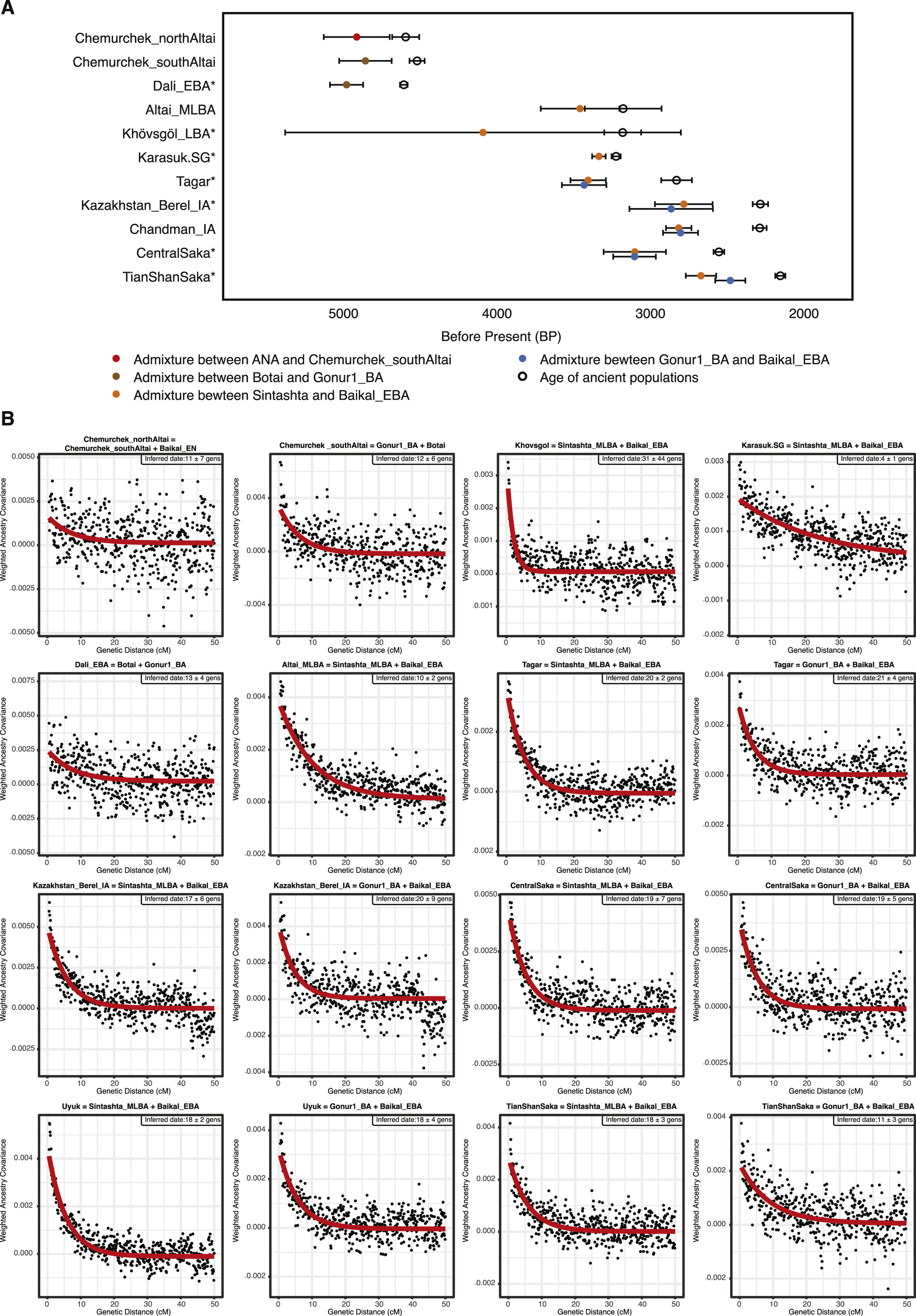
图S6 史前个体的混合年代测定,与STAR方法相关
(A) 史前个体的混合年代测定。我们使用DATES程序估计混合日期,并通过添加每个古代人群的年龄(校准后14C测年95%置信区间中心的平均值)并假设每代29年进行转换。与混合日期(彩色圆圈)相关的水平条是通过对DATES估计使用留一染色体法(leave-one-chromosome-out jackknifing)的方差和14C日期估计的方差求和的平方根来估计的,假设这两个量是独立的。已发表的群体用星号(*)标记。对于北阿尔泰切穆尔切克(Chemurchek_northAltai),我们使用贝加尔_新石器时代早期(Baikal_EN)作为ANA血统的代表来测定混合事件的日期,考虑到贝加尔_新石器时代早期样本量较大。
(B) 史前个体的血统协方差。我们展示了从DATES计算的加权血统协方差(y轴),它预计将沿遗传距离(x轴)呈指数衰减,衰减率表示自混合以来的时间,以及拟合的指数曲线(显示为红线)。我们从遗传距离0.45厘摩根开始拟合,并通过加权区块留一法(在每次运行中删除一条染色体)来估计标准误差。
尽管基于少量基因组,我们发现阿法纳谢沃人和切穆尔切克人都没有在随后的中晚青铜时代(MLBA)留下持久的基因痕迹。这与欧洲的情况明显不同,在欧洲,迁徙的早期青铜时代(EBA)草原牧民对当地人口产生了变革性的持久基因影响(Allentoft等,2015;Haak等,2015;Mathieson等,2018)。在东部草原,早期青铜时代牧民的短暂基因影响与他们强大且持久的文化和经济影响形成鲜明对比,因为早期青铜时代牧民首先引入的文化特征,如丧葬土墩建筑和乳制品畜牧业,一直延续至今。
青铜时代三重基因结构的出现之前,我们报告了早期青铜时代西贝加尔湖狩猎采集者(Baikal_EBA)和蒙古北部晚期青铜时代(LBA)牧民(Khövsgöl_LBA)之间共享的基因特征(Jeong等,2018)。这种基因特征分别由主要的ANA和次要的ANE血统成分组成,同时也与本研究分析的更早期的东贝加尔湖(Fofonovo_EN)和蒙古(centralMongolia_preBA)群体共享(图3A、3B和4A),表明这种基因特征在该地区持续存在了近三千年。以蒙古北部为中心的这种基因特征与其他青铜时代群体明显不同。总体而言,我们在晚期青铜时代蒙古发现了三个截然不同且地理分布有结构的基因库,其中Khövsgöl_LBA人群代表其中之一(图3B和4A)。其他两个,我们称之为"Altai_MLBA"和"Ulaanzuukh_SlabGrave",将在下文中描述。
在中晚青铜时代(公元前1900-900年),随着气候变化导致草原扩张,新的牧民文化从内部山区扩展到整个东部草原(Kindstedt和Ser-Od,2019)。这一时期还因首次出现马奶利用的区域性证据(约公元前1200年;Wilkin等,2020a)而著名,如今这一做法专门与酒精(airag)生产相关(Bat-Oyun等,2015),并且马匹使用急剧增强,包括骑马技术的出现,这大大扩展了对草原偏远地区的可达性。在阿尔泰-萨彦地区,与鹿石-赫列木墓地文化(DSKC)和其他未分类的中晚青铜时代墓葬类型(Altai_MLBA,n = 7)相关的乳制品牧民显示出了明确的遗传证据,表明他们是Khövsgöl_LBA相关血统与Sintashta相关的西部草原牧民(WSH)血统混合的结果(图3B;图S4B)。总体而言,他们在主成分分析(PCA)上形成了一条"Altai_MLBA"倾斜线,位于西部草原群体和贝加尔早期青铜时代/Khövsgöl晚期青铜时代(Baikal_EBA/Khövsgöl_LBA)群集之间(图2),他们在PC1轴上的位置根据其西部血统水平而变化(表S5C)。
这是辛塔什塔类型血统(在之前的研究中经常被称为"steppe_MLBA")在东部草原的首次出现,它与阿法纳谢沃和切穆尔切克人群中存在的早期西方血统截然不同,而是与欧洲绳纹陶文化人群以及后来的安德罗诺沃相关群体(如辛塔什塔)表现出密切亲缘关系(Allentoft等,2015)。在科布多省,属于鹿石-赫列木墓地文化(DSKC)和未分类中晚青铜时代(MLBA)群体的个体(分别为BER002和SBG001)具有相似的基因特征,最佳模型表明它们是Khövsgöl_LBA和辛塔什塔等量混合的结果(图3B;表S5C)。这一基因特征与之前研究中描述的蒙古北部一个偏离Khövsgöl_LBA集群的基因异常个体相匹配(ARS026;Jeong等,2018)。另外四个属于鹿石-赫列木墓地文化(ULI001)和未分类中晚青铜时代群体(BIL001、ULI003、ULZ001)的阿尔泰中晚青铜时代(Altai_MLBA)个体也符合这种混合模型,但混合比例各不相同(表S5C)。综合来看,Altai_MLBA倾斜线揭示了两个源人群的持续混合:一个与辛塔什塔/安德罗诺沃相关的西部草原(WSH)人群和一个由Khövsgöl_LBA代表的本地人群。据估计,这种混合发生在本研究分析的个体之前仅10±2代(约290年)前,这一发现与它们异质的血统比例相一致(图S6)。由于辛塔什塔文化(约公元前2200-1700年)与马拉战车等新型交通技术相关(Anthony,2010),这种血统特征在东部草原的出现表明,提高的机动能力在连接欧亚草原不同人群方面发挥了重要作用(Honeychurch,2015)。
我们数据集中的三个中晚青铜时代(MLBA)个体呈现出无法完全由Altai_MLBA倾斜线解释的基因谱系。这三个个体,包括两个阿尔泰个体(UAA001和KHI001)以及来自科布多省的UUS001,更适合以Gonur1_BA作为第三个血统来源的小贡献模型(表S5C)。综合来看,尽管阿尔泰和蒙古北部的主要中晚青铜时代墓葬传统(蒙赫海尔汗、鹿石-赫列木墓地文化和未分类中晚青铜时代)之间可能存在文化差异,但它们并不形成截然不同的基因群体。
构成异质性阿尔泰中晚青铜时代(Altai_MLBA)倾斜线的人群在阿尔泰-萨彦地区留下了后裔,我们后来在早期铁器时代(EIA)期间于蒙古西北部查德曼山的Sagly/Uyuk遗址("Chandman_IA",约公元前400-200年)中识别出这些后裔。九个Chandman_IA个体在主成分分析(PCA)上形成了一个紧密的集群,位于先前Altai_MLBA倾斜线的末端,远离Khövsgöl_LBA集群(图2)。在早期铁器时代,Sagly/Uyuk人是牧民和粟农牧民,主要分布在今天图瓦地区的上叶尼塞河地区。他们与阿尔泰地区的Pazyryk人和哈萨克斯坦东部的Saka人一起,构成了一个更广泛的斯基泰文化现象的一部分,该文化现象横跨西部草原、塔里木盆地和上叶尼塞河地区(Parzinger, 2006)。
我们发现早期铁器时代斯基泰人群系统性地偏离了早期阿尔泰中晚青铜时代倾斜线,需要引入第三个祖源成分(图3C和图4A;图S4C)。这种与中亚(高加索/伊朗高原/中亚河中地区)人群相关的血统,包括巴克特里亚-马尔吉安纳考古复合体(BMAC)(Narasimhan等,2019),在铁器时代群体中如中部萨卡人、天山萨卡人、塔加尔人(Damgaard等,2018b)和Chandman_IA中明显可见,而在早期的鹿石-赫列木墓地文化(DSKC)和卡拉苏克群体中则不存在(表S5C-S5E)。这第三个成分在这些铁器时代群体的血统中占6%-24%,而Chandman_IA中混合的日期估计在约18±4代之前,大约公元前750年,这晚于BMAC约公元前1600年的崩溃,略早于波斯阿契美尼德帝国约公元前550年的形成(图S6)。我们认为,这种伊朗相关的基因流入是由于在晚青铜时代到早期铁器时代过渡期间,与中亚河中地区(图兰)和费尔干纳地区的农牧民人口接触和混合增加所促成的。骑马在公元前第二个千年末和第一个千年初的广泛兴起(Drews,2004),以及此后马匹运输技术的不断完善,可能促进了人口接触增加和这种伊朗相关血统向草原的传播。我们的结果并不排除其他接触领域,如沿内亚山脉走廊流动性的增加,这也可能从青铜时代开始通过新疆将这种血统引入阿尔泰(Frachetti,2012)。
与阿尔泰和蒙古北部的中晚青铜时代和早期铁器时代文化相比,在蒙古东部和南部地区发现了不同的埋葬传统(Honeychurch, 2015),尤其是晚青铜时代的乌兰祖赫文化(公元前1450-1150年)和早期铁器时代的石板墓文化(公元前1000-300年)。与东部草原其他同时期人群不同,我们发现与这些埋葬类型相关的个体显示出明显的东北亚(ANA相关)基因特征,缺乏ANE和WSH混合成分(图2、3C和4)。这两个群体都是反刍动物牧民,而早期铁器时代的石板墓文化还饲养马匹(Wilkin et al., 2020a)。
乌兰祖赫和石板墓个体的基因特征在遗传上无法区分(图2;表S5C),这与考古学假设一致,即石板墓传统源自乌兰祖赫文化(Honeychurch, 2015;Khatanbaatar, 2019)。这两个群体与更早期的东蒙古青铜时代前个体(约公元前4600年)也无法区分,表明这个史前东蒙古基因库具有长期(>4,000年)的稳定性(表S5C)。在后续分析中,我们将乌兰祖赫和石板墓合并为一个单一的遗传群体("Ulaanzuukh_SlabGrave")。
Ulaanzuukh_SlabGrave遗传群簇很可能是此前描述的来自库苏古尔省的DSKC东部异常个体(ARS017)的来源(Jeong et al., 2018),以及一个来自蒙古中部的文化上未分类的个体(TSI001),该个体可追溯到晚青铜时代-早期铁器时代过渡期(图2、3B和3C;表S5C)。此外,来自蒙古西北部的蒙赫海尔汗个体KHU001除了其贝加尔早期青铜时代血统外,还具有不可忽视的Ulaanzuukh_SlabGrave血统(图S4B;表S5C)。
虽然这三个个体证明了晚青铜时代期间蒙古西北部和东部之间偶尔存在远距离接触,但我们在阿尔泰地区没有发现Ulaanzuukh_SlabGrave血统的证据,而且在中晚青铜时代期间,蒙古东部和南部以外地区的Ulaanzuukh_SlabGrave基因特征的总体频率非常低。在早期铁器时代,石板墓文化向北扩张,有时会破坏和连根拔起其路径上的前DSKC墓葬(Fitzhugh, 2009;Honeychurch, 2015;Tsybiktarov, 2003;Volkov, 2002),并最终到达了远至东部贝加尔地区,这反映在本研究中石板墓个体PTO001的基因特征中(图3C)。
总体而言,我们的发现揭示了青铜时代东部草原人群在早期铁器时代末期前存在明显的东西方基因分化。来自蒙古中部和南部的进一步取样将有助于完善这些血统特征的空间分布,以及我们当前发现的代表性。
匈奴帝国,第一个草原游牧帝国的崛起
从东部草原的史前人口中,大规模的政治实体开始于公元前一千年末期发展。匈奴是第一个有历史记载的由游牧民族建立的帝国,其成立被认为是东部草原社会政治历史的分水岭事件(Brosseder and Miller, 2011; Honeychurch, 2015)。匈奴从公元前三世纪到公元一世纪在东亚和中亚保持着政治优势。构成匈奴帝国的人民的文化、语言和遗传构成一直是研究的重点,同时他们与东部草原上其他同时代和后续游牧群体的关系也备受关注。
在此,我们报告了来自蒙古各地60个匈奴时期个体的全基因组数据,这些个体的年代约为公元前200年至公元100年,因此跨越了整个匈奴帝国时期。尽管大多数个体属于晚期匈奴时期(公元前50年后),但有13个个体早于公元前100年,其中包括来自北部早期匈奴边疆遗址Salkhityn Am(SKT)和Atsyn Gol(AST)的12个个体,以及来自蒙古东部早期匈奴遗址Jargalantyn Am(JAG)的1个个体。
我们观察到两个独特的人口学过程促成了早期匈奴的形成。首先,半数早期个体(n = 6)形成一个遗传集群(earlyXiongnu_west),与来自阿尔泰-萨彦地区的前身萨格雷/乌尤克文化的Chandman_IA相似(图2)。他们的血统92%来自Chandman_IA,其余部分归因于额外的伊朗相关血统,我们使用BMAC作为代理模型(图3D和4D;表S5F)。这表明在早期铁器时代Chandman_IA萨格雷/乌尤克文化中发现的低水平伊朗相关基因流可能在公元前第一千年后半期继续存在,并传播到蒙古西部和北部。其次,六个个体("earlyXiongnu_rest")在earlyXiongnu_west和Ulaanzuukh_SlabGrave集群之间呈中间状态;四个携带不同程度的earlyXiongnu_west(39%-75%)和Ulaanzuukh_SlabGrave(25%-61%)相关血统,而两个(SKT004,JAG001)与Ulaanzuukh_SlabGrave集群无法区分(图3D;表S5F和S5G)。这种连接earlyXiongnu_west和Ulaanzuukh_SlabGrave基因库的遗传连续体标志着东部草原上两个深度分歧且截然不同的血统的统一——在西部的DSKC、蒙赫海尔汗和萨格雷/乌尤克文化的后裔与东部的乌兰祖赫和石板墓文化的后裔之间。总体而言,从先前的萨格雷/乌尤克文化继续的低水平伊朗相关基因流入以及突然出现的将东部草原基因库联合起来的新型东西方混合是与匈奴崛起相关的两个决定性人口学过程。
在晚期匈奴个体中,我们发现更高的基因异质性(图2),他们在主成分分析图上的分布表明,早期匈奴中明显的两种人口学过程在晚期匈奴时期继续存在,但同时还增加了新的基因流动浪潮和复杂方向。在47个晚期匈奴个体中,一半(n = 26)可以通过与早期匈奴相同的混合过程进行充分建模:22个作为Chandman_IA+Ulaanzuukh_SlabGrave的混合,2个(NAI002,TUK002)作为Chandman_IA+BMAC或Chandman_IA+Ulaanzuukh_SlabGrave+BMAC的混合,以及2个(TUK003,TAK001)作为earlyXiongnu_west+Ulaanzuukh_SlabGrave或earlyXiongnu_west+Khovsgol_LBA的混合(图3D和4D;表S5G)。另外两个个体(TEV002,BUR001)也可能源自早期匈奴基因池,尽管其模型的p值略低于0.05阈值(表S5G)。然而,另外11个在PC1上具有最高西欧亚亲缘性比例的晚期匈奴个体无法使用BMAC或任何其他古代伊朗相关人群进行建模。相反,他们与来自西部和中部草原各地的古代萨尔马特人群聚在一起(图2)。
混合模型建模证实了晚期匈奴中存在萨尔马特相关的基因库:三个个体(UGU010、TMI001、BUR003)与萨尔马特人在基因上无法区分,两个个体(DUU001、BUR002)是萨尔马特和BMAC的混合,三个个体(UGU005、UGU006、BRL002)是萨尔马特和乌兰祖赫_石板墓的混合,三个个体(NAI001、BUR004、HUD001)的基因模型需要同时包含萨尔马特、BMAC和乌兰祖赫_石板墓的成分(图3D;图S4D;表S5G)。此外,在PC1轴上显示最高东欧亚亲缘性的八个个体与乌兰祖赫_石板墓和Khövsgöl_LBA两种基因谱系都不同,沿着PC2轴表现出与更南方的现代东亚人群的亲缘性(图2)。其中六个个体(EME002、ATS001、BAM001、SON001、TUH001、YUR001)可以很好地被建模为乌兰祖赫_石板墓和汉人的混合体(表S5F和S5G),其中YUR001特别表现出与先前发表的两名汉帝国士兵(Damgaard等,2018b)密切的基因相似性,我们将这种基因谱系称为"Han_2000BP"(表S5G)。剩下的两个个体(BRU001、TUH002)相似但还需要添加萨尔马特血统(表S5G)。因此,晚期匈奴的特点是两个额外的人口学过程,这使他们与早期匈奴区分开来:一是来自新的萨尔马特相关西方血统源的基因流,二是与同时期中国汉帝国人民的交流与混合加强。Egyin Gol匈奴墓地的先前研究报告了来自西方和东方欧亚起源的线粒体单倍群(Keyser-Tracqui等,2003),这与我们从全基因组数据中发现的东西方混合结果一致。总的来说,这些结果与历史记录很好地吻合,这些记录记载了匈奴对其邻国的政治影响,包括中亚丝绸之路王国和汉朝中国,以及蒙古地区人口的迁入和迁出(Miller,2014)。总体而言,匈奴时期可以被描述为一个大规模且广泛的基因流动时期,它始于统一蒙古西部和东部的基因库,最终统一了西亚和东亚的基因库。
匈奴后政权中的基因异质性波动
在匈奴帝国于公元100年左右崩溃后,在接下来的几个世纪里,一系列游牧政权在政治上分裂的东部草原地区兴衰更替:鲜卑(约公元100-250年),柔然(约公元300-550年),突厥(公元552-742年)和回鹘(公元744-840年)。尽管我们对早期中世纪时期的样本代表性不均衡,包括1个归属于鲜卑或柔然时期的未分类个体(TUK001),8个来自突厥墓葬环境的个体,以及13个来自回鹘墓地的个体,但很明显这些个体的基因特征与前一个匈奴时期不同,表明此时有新的基因流入蒙古,使他们沿PC3轴发生位移(图2)。个体TUK001(公元250-383年校准年代),其墓葬是对早期匈奴墓地的侵入,具有最高的西欧亚亲缘性。这种血统与萨尔马特人不同,更接近于具有BMAC/伊朗相关血统的古代人群(图2)。在具有最高东欧亚亲缘性的个体中,两个突厥时期的个体和一个回鹘时期的个体(ZAA004,ZAA002,OLN001.B)与乌兰祖赫_石板墓群体在基因上无法区分。另一个个体(TUM001),从唐朝时期突厥精英使者墓道中发现,具有高比例的汉族相关血统(78%;图3E和图4B;图S4E;表S5H)。这名男性与两只狗一起埋葬,可能是被牺牲以守卫墓道入口的中国随从(Ochir等,2013)。其余17个突厥和回鹘个体显示出中间基因特征(图3E)。
早期中世纪时期的高度基因异质性鲜明地体现在回鹘时期的奥伦多夫墓地(OLN;图2)的12个个体身上,该墓地位于回鹘首都奥尔杜-巴利克附近。其中6个个体来自同一座墓葬(19号墓),其中只有两个人有亲缘关系(OLN002和OLN003,二级亲属;表S2D);没有更近亲缘关系的事实引发了对此类墓葬功能及其中埋葬者社会关系的疑问。大多数回鹘时期个体表现出高度但可变的西欧亚血统——最佳模型为阿兰人(一个历史上的游牧牧民群体,可能是萨尔马特人的后裔,与匈人同时代(Bachrach, 1973))和伊朗相关(BMAC相关)血统的混合——以及乌兰祖赫_石板墓(ANA相关)血统(图3E)。本研究中古代突厥和回鹘个体的混合日期估计约为公元500年:突厥个体之前8±2代,回鹘个体(由ZAA001和奥伦多夫个体代表)之前12±2代。
蒙古帝国的兴起
在回鹘帝国于九世纪中叶灭亡后,中国东北的契丹人于公元916年建立了强大的辽朝。契丹人控制了东部草原的大片区域,据记载他们在其征服的领土内迁徙人口(Kradin和Ivliev,2008),但在蒙古境内发现的契丹时期墓地很少。我们的研究包括来自布尔干省的三个契丹个体(ZAA003、ZAA005、ULA001),他们都具有明显的东欧亚基因特征(图2),西欧亚血统不到10%(图3F和4B;表S5I)。这可能反映了说蒙古语的契丹人的东北亚起源,但需要更大的样本量来充分描述蒙古境内契丹人群的基因特征。公元1125年,契丹帝国被女真人的金朝所灭,而金朝又于公元1234年被蒙古人征服。
蒙古帝国的鼎盛时期
在其最大范围内,蒙古帝国(公元1206-1368年)横跨了欧亚大陆近三分之二的土地。它是世界上最大的连续陆地帝国,这个国际化的实体由流入草原腹地的多样化人口组成。我们分析了62位蒙古时期的个体,他们的埋葬方式与当地低级精英的埋葬一致。研究中没有包括王室或区域精英的墓葬,也没有包括来自国际化首都哈拉和林的个体。虽然我们发现蒙古时期的个体具有多样性,但他们表现出比匈奴时期个体低得多的基因异质性(图2),而且他们几乎完全缺乏曾在匈奴和早期北部/西部MLBA文化中存在的残留ANE相关血统(以Chandman_IA和Khövsgöl_LBA的形式)。平均而言,蒙古时期的个体比先前帝国具有更高的东欧亚亲缘性,这一时期标志着现代蒙古基因库形成的开始。我们发现大多数历史上的蒙古人可以通过以下祖源代表的三方混合模型很好地拟合:乌兰祖赫_石板墓、汉族和阿兰人。与其PCA位置一致(图2),蒙古时期的个体作为一个群体可以用仅15%-18%的西部草原血统(阿兰人或萨尔马特人)建模,但需要55%-64%的乌兰祖赫_石板墓和21%-27%的汉族相关血统(表S5I)。将相同的模型分别应用于每个个体,这个三源模型充分解释了61个古代蒙古人中的56个(基于p值阈值0.05),以及一个未分类的晚期中世纪个体(SHU002),其年代约为蒙古帝国初期(表S5J)。
自蒙古帝国于公元1368年覆灭以来,蒙古人口的基因特征并未发生实质性变化。蒙古帝国时期形成的基因结构继续特征性地存在于当今居住在蒙古和俄罗斯的蒙古语系人群中。我们使用基于个体的qpWave分析,研究了历史上的蒙古人与七个现代蒙古语系群体(蒙古人、卡尔梅克人、布里亚特人、哈穆尼干人、达斡尔人、土族人和蒙古拉人)之间的基因谱系关系。在当前数据的分辨率下,61位历史蒙古人中有34位在基因上与至少一个现代蒙古语系人群谱系相同(图S7B)。蒙古帝国对东部草原的政治和基因景观的重构产生了深远影响,这些影响在帝国衰落后仍然长期存在,至今在蒙古仍然明显可见。
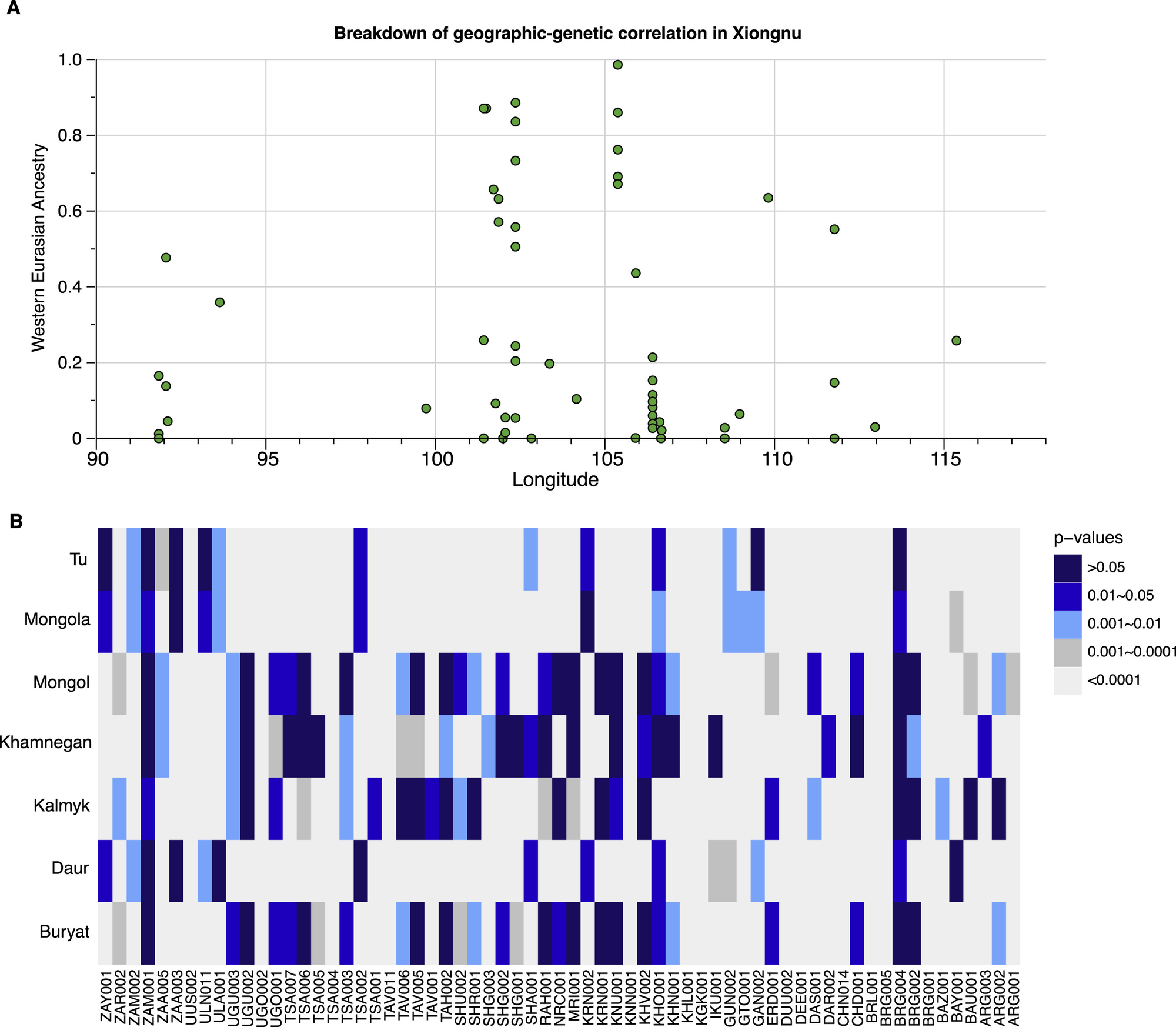
图S7 匈奴地理和遗传分布分析以及蒙古时期与现代人群比较,与图3和STAR方法相关
(A) 匈奴时期地理-遗传相关性的分析。我们展示了匈奴时期所有个体/群体的西欧亚血统比例(y轴)与其考古遗址经度(x轴)的关系。使用萨尔马特人作为西欧亚来源的模型的个体估计原始数据可在表S5G中找到。与中晚青铜时代/早期铁器时代(MLBA/EIA)个体不同(图3),来自更西部遗址的匈奴个体并没有比东部遗址个体具有更高比例的西欧亚血统。
(B) 使用qpWave比较古代蒙古个体与七个现代蒙古语系人群间的遗传同质性。我们报告了使用人类起源数据集中的七个现代蒙古语系人群(布里亚特人、达斡尔人、卡尔梅克人、哈穆尼干人、蒙古人、蒙古拉人和土族人)的每个基于个体的qpWave {古代蒙古个体;蒙古语系群体}的p值。当qpWave的p值>0.05时,表明y轴上的古代个体在遗传上与x轴上显示的现代蒙古语系人群无法区分。较小的p值表明古代个体与现代群体存在显著差异。
东部草原周期性混合的功能和性别方面
为了研究东部草原周期性混合的功能方面,我们估计了与乳糖消化(LCT/MCM6)、牙齿形态(EDAR)、色素沉着(OCA2、SLC24A5)和酒精代谢(ADH1B)相关的五个SNPs的群体等位基因频率(图5A)。首先,我们发现尽管有游牧生活方式和广泛的牛奶消费直接证据(Jeong等,2018;Wilkin等,2020a),东部草原的中晚青铜时代和早期铁器时代个体并没有任何导致乳糖酶持续性的衍生突变。随后时期的个体确实具有今天在欧洲广泛存在的衍生突变(rs4988235),但频率极低(约5%),且随时间推移频率没有增加(图5A)。这相当引人注目,因为除了其他乳制品外,一些当代蒙古牧民在夏季每天消费高达4-10升airag(发酵马奶,约2.5%乳糖)(Bat-Oyun等,2015),导致每日摄入100-250克乳糖。叶尼塞流域早期铁器时代的岩画已描绘了airag的生产(Dėvlet,1976),历史蒙古人的记录显示大量频繁消费airag,以及各种额外的液态和固态反刍动物乳制品(Bayarsaikhan,2016;Onon,2005),这也得到了古代蛋白质组学证据的确认(Jeong等,2018;Wilkin等,2020a)。蒙古人在缺乏乳糖酶持续性的情况下如何能够消化数千年如此大量的乳糖尚不清楚,但可能与他们据报道不寻常的肠道微生物组结构有关,现今这种结构高度富含乳糖消化Bifidobacterium菌种(Liu等,2016)。
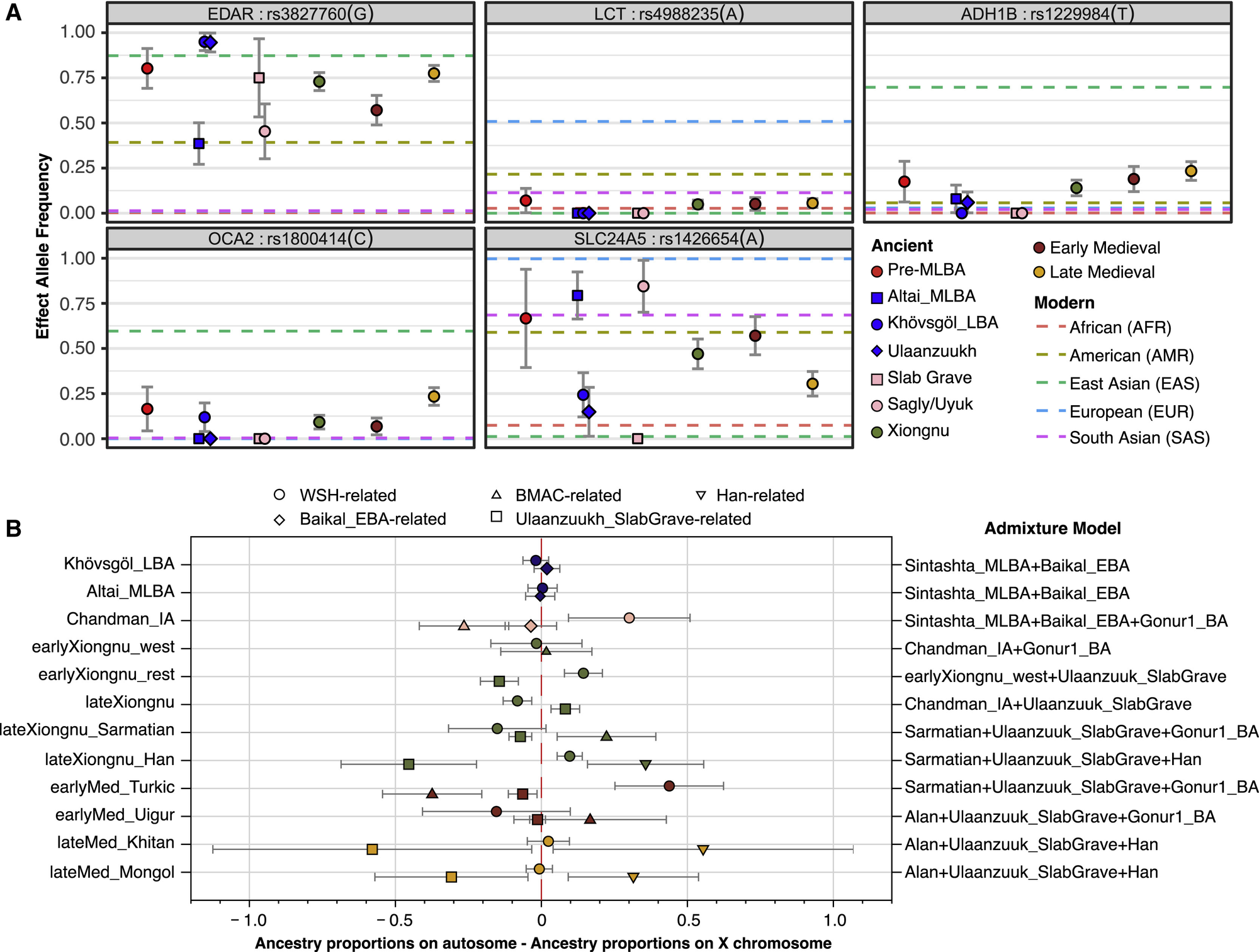
图5 功能等位基因频率和性别偏向的基因混合模式
(A) 五种表型SNP随时间变化的等位基因频率。对于有效等位基因,我们展示了每个古代群体的最大似然频率估计和一个标准误差条。前MLBA类别对应于蒙赫海尔汗之前所有古代群体的总和。匈奴、早期中世纪和晚期中世纪分别对应于每个时期所有古代群体的总和。水平虚线显示来自1000基因组计划的五个超级人群的等位基因频率信息。
(B) 按时期和人群划分的性别偏向基因混合模式。我们为每个具有WSH/伊朗/汉相关祖源基因混合的古代个体计算了Z值。正值表明常染色体上有更多的WSH/伊朗/汉相关祖源,即男性驱动的混合。查看图S2C获取个体Z值。
经历区域性选择性扫荡的遗传标记表现出的等位基因频率变化与全基因组祖源谱系的变化相关(图5A)。例如,EDAR(外胚层发育不良蛋白A受体)基因中的rs3827760和SLC24A5(溶质载体家族24成员5)基因中的rs1426654分别是东亚人和西欧亚人群中众所周知的正向选择目标(Sabeti等,2007)。我们的MLBA和EIA人群在这两个SNPs的等位基因频率上显示出强烈的人群分化:rs3827760频率在具有较高东欧亚亲缘性的群体(Khovsgol_LBA,Ulaanzuukh_SlabGrave)中明显更高,而rs1426654在Altai_MLBA和Chandman_IA中更高(表S2E)。我们发现在东亚人群中经历了更近期正向选择(Donnelly等,2012;Li等,2011)的两个SNPs:ADH1B(醛脱氢酶1B)基因中的rs1229984和OCA2(眼皮肤白化病II型)基因中的rs1800414,在MLBA和EIA时期要么不存在,要么频率极低(当时东欧亚祖源主要与ANA相关),但随着时间推移,通过与中国帝国和其他群体的互动,东亚祖源比例增加,这些SNPs的频率也随之增加(表S2E)。
最后,我们研究了东部草原人口历史的性别维度。性别偏向的基因混合模式可以为迁徙、社会亲属关系和家庭结构的性别方面提供信息。我们在EIA Sagly/Uyuk人群和突厥时期观察到明显的男性偏向WSH混合信号(即更多正向Z值;图5B),这也与Y染色体谱系Q1a的减少以及西欧亚谱系如R和J的同时兴起相对应(图S2A)。在后期的契丹和蒙古帝国时期,我们观察到东亚相关祖源的显著男性偏向(图S2C),这也可以从Y染色体谱系O2a频率的上升中看出(图S2A)。匈奴时期展现了最复杂的男性偏向混合模式,不同的人口基因亚群显示出不同来源的男性偏向混合证据(图S2C)。
在匈奴人中,我们还检测到10对基因亲缘关系,包括一对埋葬在Jargalantyn Am同一墓穴中的父女(JAG001和JAA001),以及Il'movaya Pad的一对母子(IMA002和IMA005),Tamiryn Ulaan Khoshuu的一对兄妹(TMI001和BUR003),和Salkhityn Am的一对兄弟(SKT002和SKT006)(表S2D)。在剩余的六对中,有三对是埋葬在同一地点的女性-女性亲缘关系,表明匈奴群体中存在扩展的女性亲属关系。基于常染色体短串联重复序列(STR)数据的先前研究也报告了在Egyin Gol匈奴墓地中单一地点内存在的一级亲属关系(Keyser-Tracqui等,2003)。这些关系结合墓葬特征,为我们提供了了解匈奴帝国内部地方血统和亲属结构的首批线索,而这些结构在其他方面仍知之甚少。
讨论
东部草原的人口历史以多样化的东西欧亚基因库的反复混合为特征。然而,这并非简单的迁徙浪潮,东部草原的人口事件一直是复杂且多变的。通过生成超过200个全基因组古代数据集,我们提供了这一动态人口历史的首个基因证据,时间跨度从约公元前4600年直至蒙古帝国末期。我们发现,在全新世中期,东部草原由具有ANA和ANE祖源的狩猎采集者占据,随后在青铜时代转变为乳制品游牧经济。装备有车辆和家畜的迁徙亚姆纳亚/阿法纳西耶沃草原牧民(Kovalev和Erdenebaatar,2009年),似乎在约公元前3000年首次引入了反刍动物乳制品游牧业(Wilkin等,2020a),但出人意料的是,他们几乎没有留下持久的基因影响,这与欧洲的情况不同(Allentoft等,2015年;Haak等,2015年;Mathieson等,2015年)。到了中晚青铜时代(MLBA),无论祖源如何,东部草原的人口都已采用反刍动物乳制品游牧业(Wilkin等,2020a),这种生存方式一直延续至今,后期又增加了晚青铜时代的马奶和蒙古时期的骆驼奶(Wilkin等,2020a;Bat-Oyun等,2015年;Kindstedt和Ser-Od,2019年)。然而令人费解的是,尽管这一遗传特征通过后续西方群体的多次迁徙而被引入,但在这5,000年的历史中并没有发现对乳糖酶持续性的选择证据。这表明亚洲地区存在一种不同的乳糖适应轨迹,至今仍未得到解释。
在MLBA(中晚青铜时代)期间,我们观察到东部草原形成了一个三重基因结构,其特点是东部继续保持青铜时代前的ANA祖源,北部呈现青铜时代前ANA-ANE祖源之间的基因变异梯度,而西部则出现了新的辛塔什塔相关WSH祖源比例增加的现象。辛塔什塔,一种与欧洲绳纹陶文化有基因联系的西部森林草原文化(Mathieson等,2015),是青铜冶金和战车技术的大师(Anthony,2010),这种祖源在东部草原的出现可能与新技术(尤其是与马相关的技术)的引入有关。DSKC(鹿石-赫列克苏尔文化)遗址特别显示了马匹用于运输甚至可能骑乘的广泛证据(Taylor等,2015),基因分析已证明这些动物与辛塔什塔战车马之间存在密切联系(Fages等,2019)。青铜时代东部草原人口之间的这种强烈的东西方基因分化持续了一千多年,直至早期铁器时代(EIA)结束,当时出现了广泛骑马的第一个明确证据(Drews,2004),某些群体,特别是东部板石墓文化(Honeychurch,2015)的流动性增强,开始打破这种结构。最终,三种主要祖源相遇并混合,这与匈奴帝国的崛起同时发生。匈奴人以极高的基因异质性和多样性增加为特征,来自中国、中亚和西部草原(萨尔马特相关)的新增祖源迅速进入基因库。
早期中世纪时期的基因数据相对稀疏且不均衡,在匈奴与突厥时期之间400年的间隔中,几乎没有鲜卑或柔然遗址被确认。我们观察到在突厥和回鹘时期存在高度的基因异质性和多样性,而在回鹘帝国崩溃后,我们记录到晚中世纪时期发生了最后一次主要的基因转变,朝着更大的东欧亚血统方向发展,这与历史文献记载的通古斯语系(女真)和蒙古语系(契丹和蒙古)群体从东北向东部草原的扩张相一致(Biran, 2012)。我们还观察到,这种东亚相关血统更多是通过男性祖先而非女性祖先带入晚中世纪人口的。到蒙古时期结束时,东部草原的基因构成已经发生了戏剧性的变化,几乎不再保留曾在其史前时期占主导地位的ANE祖源。如今,ANE祖源仅在孤立的西伯利亚群体和美洲原住民中以相当数量存活(Jeong et al., 2019)。历史上蒙古人的基因特征仍然反映在当代蒙古人身上,表明这一基因库在过去约700年中相对稳定。
已经记录了东部草原的几个关键基因转变时期,未来的研究可能能够探索这些转变是否也与文化和技术创新相关联,以及这些创新如何影响政治格局。将这些发现与马匹技术和牧业实践变化的研究结合起来,以及牲畜特征和品种的变化,可能会特别有启发性。本研究代表了东部欧亚草原第一个大规模的古基因组研究,它揭示了该地区极其复杂和动态的基因多样性。尽管取得了这些进展,在中亚和东亚,特别是中国东北、塔里木盆地和哈萨克东部草原等地区,仍然需要进一步的基因研究,以全面揭示欧亚草原的人口历史及其在世界史前史中的关键作用。
翻译:蒙古_科布多_中晚青铜时代至匈奴晚期早期中世纪_o:khi001.ag-KHI001
翻译:人类考古骨骼材料,本研究,KHI001(AT-398)
翻译:新的基因群体:阿尔泰_中晚青铜时代(7个样本)、乌兰佐克_板石墓文化(11/16个样本)、UAA001(1个样本)、KHI001(1个样本)、UUS001(1个样本)、KHU001(1个样本)和TSI001(1个样本)
我们还确定了三个不属于上述三个基因簇的离群个体。来自阿尔泰的UAA001(Mönkhkhairkhan)与使用Afanasievo、Baikal_EBA和Gonur1_BA的三向混合模型吻合良好(表S5C),尽管他们的年代比Afanasievo文化晚约1500年。来自阿尔泰的KHI001(未分类文化)与使用Sintatash、Baikal_EBA和Gonur1_BA的三向混合模型吻合良好(p值 = 0.056;表S5C),表现出来自Gonur1_BA的微小基因成分。另外,KHI001也可以被建模为Afanasievo和Khövsgöl_LBA之间的双向混合(p值 = 0.117;表S5C);然而,这个模型的优先级低于前一个模型。来自科布多省的UUS001(DSKC)与使用Sintashta、eastMongolia_preBA和Gonur1_BA的三向模型吻合良好(表S5C)。鉴于UUS001和eastMongolia_preBA个体之间的时间差异(约3,000年),UUS001的混合伙伴更可能与乌兰佐克簇有关;乌兰佐克与eastMongolia_preBA共享高度的血统,且与UUS001个体同时代,一些乌兰佐克个体在主成分分析中与eastMongolia_preBA个体(SOU001)非常接近。
人类考古学骨骼材料,即人类骨考古学遗存,是从考古遗址中发掘出的骨骼和牙齿。这些遗存为了解过去人群的生活提供了宝贵的见解,提供有关他们的健康、饮食、生活方式甚至文化习俗的信息。人类骨考古学家使用各种技术对这些遗存进行研究,以了解过去生活的人们。
从考古骨骼材料中可以了解什么?
- **人口统计学:**可以从骨骼遗存中确定年龄、性别、身高和人口历史。
- **健康与疾病:**骨骼证据可以揭示伤害、疾病(如关节炎或感染)和营养不良。
- **生活方式和活动:**骨骼标记可以指示体力活动水平、劳动类型和其他生活方式因素。
- **饮食:**骨骼的同位素分析可以揭示有关过去饮食的信息,包括所消费的植物和动物性食物的比例。
- **文化习俗:**可以识别出埋葬习俗、食人证据或出于象征目的对骨骼遗存的改造。
- **迁徙和人口流动:**通过分析骨骼特征并比较不同遗址之间的差异,研究人员可以调查人口的流动。
考古骨骼遗存如何被研究?
- **视觉检查和测量:**骨考古学家仔细检查和测量骨骼和牙齿,寻找年龄、性别和健康的特定特征和指标。
- **化学和物理分析:**同位素分析、DNA分析和显微镜检查等技术有助于进一步揭示有关饮食、祖源和疾病的详细信息。
- **上下文分析:**遗存的位置和相关的文物也很重要,提供有关埋葬习俗和文化背景的见解。
研究考古骨骼遗存的重要考虑因素:
- **保存状况:**骨骼的状况可能显著影响从中获取的信息量。
- **伦理考虑:**人类骨骼遗存通常被尊重和敏感地对待,研究人员必须注意文化和宗教信仰。
- **跨学科方法:**人类骨考古学常常涉及与遗传学、考古学和人类学等其他学科的合作。
 陕公网安备61011302002223号
陕公网安备61011302002223号