全基因组序列提供对喜马拉雅人类群体形成和适应的见解
**亮点•** 喜马拉雅地区深层次的人口结构始于10,000多年前 *EPAS1*和其他适应性变异在高海拔人群中广泛存在 与东亚和中南亚的基因流动是双向的且具有时间结构 新的适应性信号与缺氧、代谢、免疫和体力活动相关 [https://www.cell.com/current-biology/fulltext/S0960-9822(25)00808-5](https://www.cell.com/current-biology/fulltext/S0960-9822(25)00808-5) **摘要**高海拔环境对人类生存和繁殖构成重大挑战,引起了人们对这些地区人口统计和适应性历史的极大关注。
亮点•
喜马拉雅地区深层次的人口结构始于10,000多年前
EPAS1和其他适应性变异在高海拔人群中广泛存在
与东亚和中南亚的基因流动是双向的且具有时间结构
新的适应性信号与缺氧、代谢、免疫和体力活动相关
https://www.cell.com/current-biology/fulltext/S0960-9822(25)00808-5
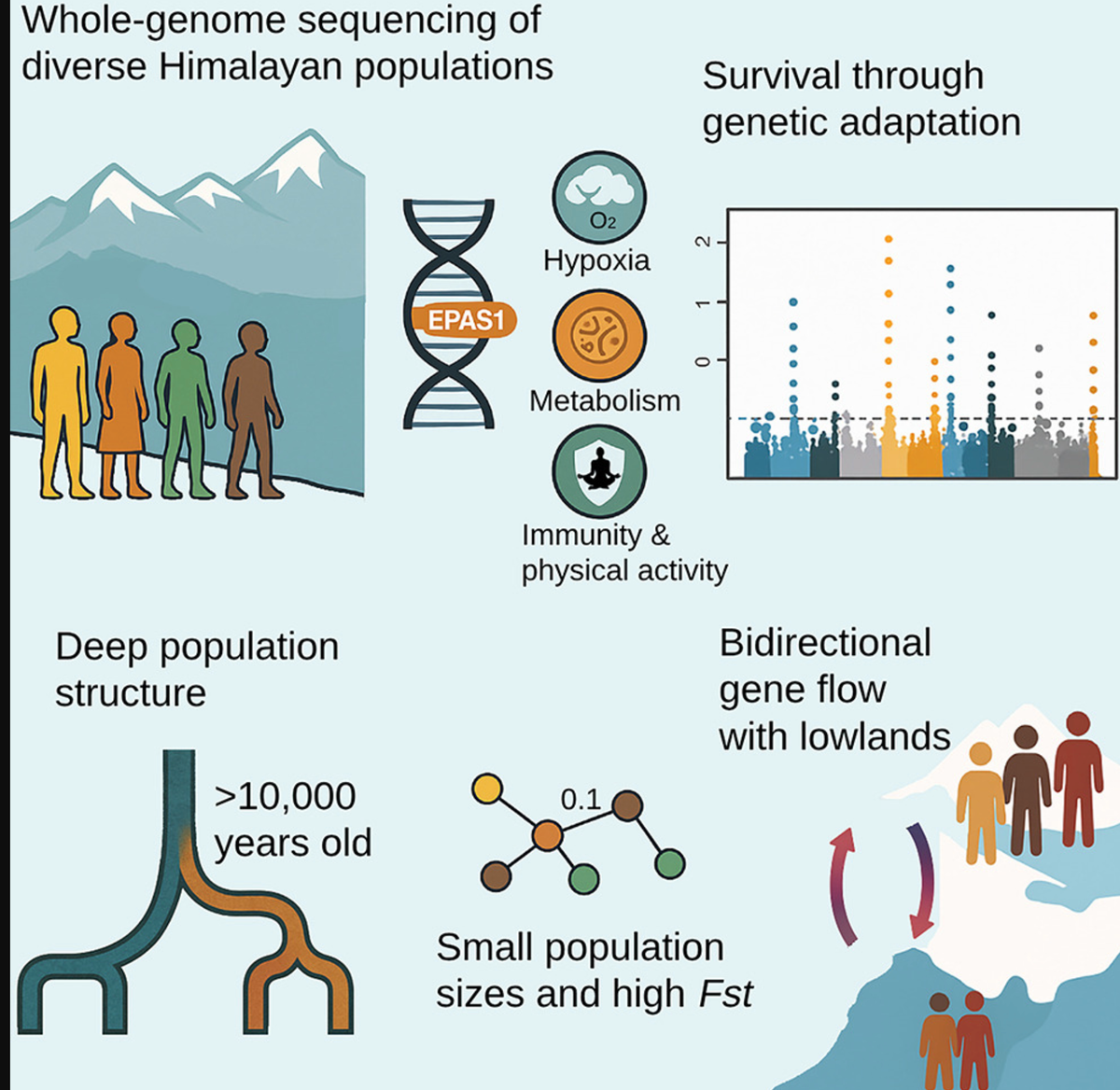
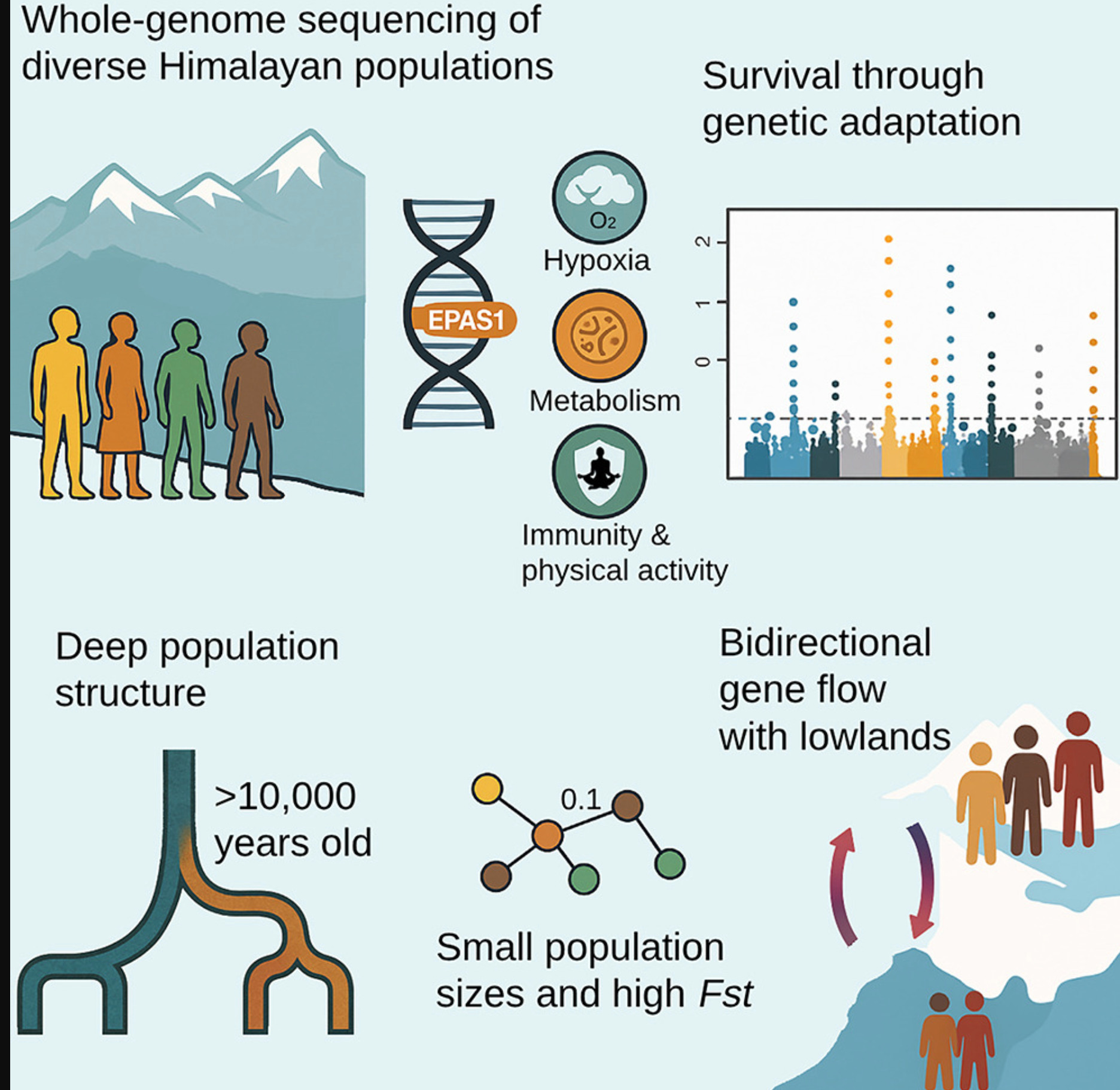
摘要高海拔环境对人类生存和繁殖构成重大挑战,引起了人们对这些地区人口统计和适应性历史的极大关注。在此,我们展示了来自不同喜马拉雅人群的全基因组序列,提供了对该地区基因组历史的新见解。我们发现喜马拉雅地区的人口结构始于约10,000年前,比考古证据显示的2,500米以上永久居住早了约6,000年。在所有高海拔人群中广泛存在的渗入适应性EPAS1单倍型突显了共同的基因起源及其对该地区生存的重要性。我们还在与缺氧、体力活动、免疫和代谢相关的基因中发现了额外的选择信号,这些都可能促进了对恶劣环境的适应。随着时间推移,不断增强的基因结构导致了今天观察到的强烈分化的民族群体的出现,其中许多群体在整个历史中保持了较小的有效人口规模或经历了严重的瓶颈效应。在6,000至3,000年前,随着农业的出现,少数单亲系血统变得占主导地位;然而,除了藏族人外,喜马拉雅地区没有观察到显著的人口增长。在近代历史中,我们检测到喜马拉雅山脉两侧的高海拔和低地群体之间的双向基因流动,这与历史区域强国,特别是藏族和北印度笈多帝国的兴起和扩张相吻合。在过去几个世纪,向喜马拉雅的迁移似乎伴随着附近地区的冲突和人口迁移,并显示出一些性别偏向。
关键词
引言
对于大多数人类来说,即使短暂地前往高海拔地区也是具有挑战性的,这主要是由于低氧浓度和寒冷温度等因素。全年生活在高海拔地区会带来更多的需求,例如寻找当地的食物来源和其他必需品。1 因此,高海拔生活已经被广泛研究,特别是在喜马拉雅地区,那里有许多人口现在永久居住在3,000米以上。关于他们的历史和适应性的见解来自化石、考古学、语言学和遗传学。已知最古老的喜马拉雅高海拔古人类化石是一个16万年前的丹尼索瓦人下颌骨,来自西藏,通过蛋白质分析鉴定。2 现代人类存在的证据可以追溯到3万至4万年前,包括在西藏海拔4,600米的尼亚德胡遗址发现的石器使用痕迹。3 然而,研究表明,海拔高达2,500米的全年定居村庄只是在5,200年前之后建立的,伴随着小米的种植,而海拔高达3,500米的永久定居则是在3,600年前之后发生的,当时来自西方的大麦和绵羊到达。4 如今,喜马拉雅高海拔人口主要讲博迪克语言,一项词汇统计学研究对博迪克语(包括塔芒语和以前被标记为"东博迪克"的语言亚群)进行了语言年代学估计,时间在4,000至3,000年前之间。5
对现代人群的基因分析,主要集中在藏族人,对比了多样化的线粒体DNA谱系与有限的Y染色体谱系,6,7强调了藏族人内部广泛的融合时间范围(7,400-29,900年前),并表明西藏最初的居住可追溯到约30,000年前,在7,000至10,000年前之间有额外的基因流动。7全基因组分析显示藏族人与东亚人整体上有基因亲缘关系,与汉族人的分离发生在约15,000-9,000年前8或44,000-58,000年前,基因流动持续到9,000年前。9这些与基于外显子组的估计不同,后者认为藏汉分离时间为2,750年前。10虽然该地区的古DNA研究仍然有限,但它们揭示了高海拔适应性等位基因的持续选择11以及复杂的藏族血统,主要来自4,000至9,000年前的北方东亚人群。12此外,来自尼泊尔安纳普尔纳保护区的样本,年代为1,250-3,150年前,海拔2,800-4,000米,显示与现代藏族人最近的基因亲缘关系,表明中高海拔地区的人口连续性。13
对高海拔适应的遗传研究发现在EPAS110基因的一个来自丹尼索瓦人渗入区域存在选择作用,14这一发现在多项研究中得到了重复证实,这些研究还指出了其他被选择的基因位点。8,9,15藏族人中丹尼索瓦人渗入片段的选择仍然是现代人类中有益古代混合的最强例证之一。然而,对非藏族人群中EPAS1或类似信号的探索一直有限。
在此,我们展示了来自16个不同喜马拉雅人群的87个新的高覆盖基因组序列。我们使用这些数据更广泛地研究喜马拉雅地区人类存在背后的人口统计学过程,解决之前的差异,并详细说明人口出现和扩张的时间线、遗传结构、与邻近人群的关系以及局部适应性。
讨论
喜马拉雅地区是多样化民族语言群体的家园,这些群体之间以及与全球人口的基因关系尚未被充分探索。喜马拉雅极端的海拔高度为人类提供了独特且具挑战性的环境,同时也形成了一道地理屏障,影响了人类在历史上的迁徙,创造了独特而复杂的基因多样性。为了理解这种多样性,我们分析了来自16个喜马拉雅人群的87个新的全基因组序列,以深入了解该地区人类群体的起源、结构和基因适应特征。
我们发现喜马拉雅人与东亚人约在10,000-12,000年前分化,这一时期正值气候变暖和世界多地新石器时代过渡期,许多社区逐渐从游牧生活方式转向定居农业和动物驯养。到全新世中期,农业已成为喜马拉雅北部和南部地区的主要生计方式,尽管某些狩猎采集群体在特定地区仍然存在。我们的结果表明,高海拔喜马拉雅人群在这一时期开始分化为今天的群体,其中一些在过去2,000年中持续分化,普遍保持较小的人口规模或经历人口瓶颈。
我们发现喜马拉雅内部存在令人惊讶的高水平群体结构,一些FST值与不同大陆之间的群体相当。这种强烈的分化可能源于长期的小有效人口规模导致的高遗传漂变,以及群体间有限的基因流动,进一步增加了基因结构。喜马拉雅人携带有限范围的单亲系血统,这可能表明小型创始人群体的快速扩张。Y染色体单倍群O2和D1的喜马拉雅亚分支以及线粒体DNA M9谱系全部在约2,000-5,000年前融合,这与村庄和农业的兴起时间吻合,这使得青藏高原在3,600年前得以永久定居。
在我们的数据集中,地理位置靠近青藏高原且有共同历史文化联系的中国彝族、纳西族和土族是与喜马拉雅人最密切相关的东亚人群。喜马拉雅地区向这些人群的显著基因流动促成了这种观察到的基因亲缘关系。东亚与喜马拉雅之间的基因流动似乎是双向的,东亚基因流动在中低海拔地区更为明显。海拔4,000米以上的人群,如夏尔巴人和拉雅普人,几乎不受这种东亚基因流动的影响。这与古DNA研究一致,表明中海拔的非藏族藏缅语使用者有着不同的基因历史,受到低地东亚人的混合影响。此外,我们在拉萨人群中检测到混合现象,包括一名有一半基因组来自低地东亚血统的个体,这可能反映了两地区之间的近期互动。
喜马拉雅向低地的基因流动向西延伸,影响了中南亚群体,如居住在巴基斯坦喀喇昆仑山脉附近山谷的布鲁什人。先前研究报告了藏族基因向附近巴尔蒂人的流动,时间为公元1341-819年。然而,我们的分析表明,布鲁什人更早接收到喜马拉雅基因流动,在公元800-450年之间,这一时间框架与藏族帝国向中亚扩张的时期一致。这一时期对该地区的文化、政治和宗教发展产生了重大影响,与我们的基因发现一致。与东亚一样,喜马拉雅与中南亚之间的基因流动是双向的。在我们的数据集中,我们观察到除夏尔巴人外的所有尼泊尔人群都显示出来自中南亚的基因流动,时间为公元700-300年,这与北印度笈多帝国的兴起和扩张一致,该帝国对尼泊尔大部分地区施加了政治、文化和贸易影响,显然也留下了基因印记。
尽管我们的发现表明喜马拉雅人在基因上与东亚人比与中南亚人更密切相关,但也有例外,如尼泊尔切特里人群。切特里人携带大量中南亚血统,我们估计这种血统源于公元1450-1100年间与尼泊尔人的混合。历史记载与我们的基因结果一致,表明切特里人在12世纪因军事入侵和不稳定而从印度迁移。此外,我们的测试表明,这次迁移涉及的印度男性明显多于女性。另一方面,对于同样在基因上接近中南亚人的塔鲁人群,我们的结果表明印度女性和男性的贡献相似。这与塔鲁人起源于逃避入侵的拉杰普特家族的历史记载相符,尽管我们的混合年代测定表明这些基因互动可能反映更早的事件。
先前研究表明,藏族人和其他喜马拉雅人携带与高海拔适应相关的丹尼索瓦人渗入单倍型。我们的结果证实了渗入的EPAS1区域在高海拔喜马拉雅人群中的广泛分布,突显了它在这一充满挑战的环境中生存的关键作用。我们估计丹尼索瓦人EPAS1簇的最近共同祖先可追溯到30,000-50,000年前,表明它可能由早期居住在该地区的狩猎采集者携带,随后随着高海拔扩张而传播。有人提出,这一单倍型在渗入后很长一段时间内保持选择中性,选择约在9,000年前才开始。这与古DNA证据一致,表明南部高原的血统可能早于4,400年,可追溯到早期本土觅食者。
我们没有发现高海拔喜马拉雅人整体上丹尼索瓦人渗入区域进一步选择的证据。然而,我们在门巴人群中发现了一个高频率的丹尼索瓦人单倍型,跨越NRM和MDC1,这些基因与基因组稳定性和淋巴细胞计数相关。这一单倍型在携带大量丹尼索瓦人血统的大洋洲人群中也很普遍。值得注意的是,该区域的一部分位于1000基因组计划(1KGP)短读测序可及性严格掩码之外,突显了未来需要通过长读测序和更广泛的采样来确定这些模式是反映漂变还是选择。
我们在2号染色体FAM178B处确定了第三个可能渗入的区域,在托托人群中频率高,在大洋洲人中也很普遍。有趣的是,这一区域先前被确定为丹尼索瓦人起源,并在印度尼西亚巴瑶人的憋气潜水中受到选择。FAM178B对低氧适应似乎很重要,调节血液pH值并防止CO₂积累。然而,与托托人的联系很谜,因为他们现在居住在印度低地。优先内婚的托托人群代表着曾经分布在西部杜尔斯各地的更大托托定居点的残余,远至提斯塔,这些定居点被人口更多的博多-科赫群体同化。除托托帕拉村外,原托托领地已被茶园吞噬。托托语属于迪马利语支,与博多-科赫语言显示出远亲关系。我们提出两种情景解释结果:(1)托托人可能起源于或经常访问附近不丹的高海拔地区,FAM178B处的渗入区域增强了他们的生存能力。这需要所有其他测试的邻近人群都失去了渗入区域。(2)另一种可能是,托托人从东南亚接收了显著的基因流动,在那里渗入变体的频率在我们数据集中的老挝和柬埔寨人等人群中约为25%。随后,当他们迁移定居在不丹山区附近时,这些变体可能由于漂变或适应而在托托人群中增至高频。值得注意的是,托托人在我们数据集中所有喜马拉雅人中展示了最高的全基因组东亚血统比例,并在主成分分析中靠近柬埔寨人。
我们独立于古代基因组的全基因组选择扫描也确定EPAS1变体具有最强信号。基因组区域延伸到相邻基因,包括TMEM247、ATP6V1E2、RHOQ、PIGF和CRIPT,这些都先前被认为是藏族人适应氧气水平变化的候选基因。另一个强信号出现在1号染色体上,跨越GNPAT、TRIM67和DISC1;后两者先前与低氧适应有关,以及附近的EGLN1。GNAPT先前与藏族人紫外线防护的皮肤色素沉着有关。我们还在4号染色体上的TBC1D1基因中确定了新的选择信号,该基因在葡萄糖和脂质利用方面起着关键作用,并影响骨骼肌中的能量底物偏好。值得注意的是,居住在高海拔地区的个体与低地地区相比,具有较低的空腹血糖和增强的葡萄糖耐受性,加上新出现的证据表明高海拔地区肥胖和糖尿病的发病率较低。此外,这个基因与鸡的耐寒适应有关,可能通过其调节能量平衡的作用。我们还在22号染色体的CLTCL1处发现了强选择信号,该基因负责指导CHC22蛋白的产生,在调节脂肪和肌肉细胞中的葡萄糖运输方面起重要作用。其他可能的适应信号包括PAX5(B细胞发育)、VTCN1和ZBTB20(T细胞调节)、SYNPO2L(心肌细胞结构/功能)、CETP(胆固醇酯/甘油三酯转移和HDL代谢)以及ASPHD2(双加氧酶和金属离子结合)。这些基因共同携带的变体在高海拔人群中与低地喜马拉雅人和邻近人群相比表现出显著分化。这些基因变异可能在塑造细胞和代谢功能方面发挥了作用,潜在地促进了在喜马拉雅具有挑战性条件下的生存。
总之,我们的研究显著推进了对喜马拉雅人群基因多样性、起源和适应性的理解。虽然先前研究主要集中在少数藏族群体上,但我们的调查引入了对该地区鲜为人知的人群和全基因组水平的见解。这一更广泛的视角增强了我们对由喜马拉雅独特环境条件塑造的复杂基因多样性和适应过程的理解。
 陕公网安备61011302002223号
陕公网安备61011302002223号