人类生态位的重大扩张早于走出非洲扩散
所有现代欧亚人的大部分祖先都可以追溯到大约5万年前(ka)从非洲迁出的一小群人口。相比之下,化石证据表明存在更早期的迁出非洲现象。这些证据只能用早期迁徙对后来的主要迁徙浪潮几乎没有基因贡献来解释。因此,一个关键问题是什么因素促成了后来成功的迁徙,使人类能够在非洲以外实现长期定居。我们在此展示,人类在非洲内部生态位宽度的显著扩张先于这次后期迁徙。我们建立了一个泛非洲的考古遗址年代测定数据库,并使用物种分布模型(SDMs)量化了过去12万年中生物气候生态位的变化。我们发现人类生态位从7万年前开始大幅扩张,这种扩张是由人类增加对从森林到干旱沙漠等多样化栖息地类型的利用所驱动的。因此,5万年后迁出非洲
摘要
所有现代欧亚人的大部分祖先都可以追溯到大约5万年前(ka)从非洲迁出的一小群人口。相比之下,化石证据表明存在更早期的迁出非洲现象。这些证据只能用早期迁徙对后来的主要迁徙浪潮几乎没有基因贡献来解释。因此,一个关键问题是什么因素促成了后来成功的迁徙,使人类能够在非洲以外实现长期定居。我们在此展示,人类在非洲内部生态位宽度的显著扩张先于这次后期迁徙。我们建立了一个泛非洲的考古遗址年代测定数据库,并使用物种分布模型(SDMs)量化了过去12万年中生物气候生态位的变化。我们发现人类生态位从7万年前开始大幅扩张,这种扩张是由人类增加对从森林到干旱沙漠等多样化栖息地类型的利用所驱动的。因此,5万年后迁出非洲的人类在面对气候挑战性栖息地时,具备了原始人类中独特的生态适应性,这为他们的适应性成功提供了一个关键机制。
遗传证据(包括与欧亚大陆古人类杂交的模式)表明,非洲以外所有现代人群的大部分祖先都来自于一次始于约5万年前的全球性扩张(参考文献1-9)。然而,化石记录显示早期也发生过人类迁徙(参考文献10-16)。这些走出非洲的迁徙很可能发生在撒哈拉-阿拉伯干旱地带的多次湿润期,最显著的是在上次间冰期,即约12.5万年前(参考文献10、17)。因此,一个重要问题是,为什么这些早期迁徙并不足以在当今非洲以外人群的基因中留下可检测的遗传痕迹,早期智人未能在非洲以外地区建立长期存活的人群。
研究人员提出了一系列解释来说明现代人类为何较晚走出非洲。一些研究认为,非洲的突然气候变化(参考文献18)以及人类认知、技术和生存方式的重大转变促进了新的"生态位拓展"创新的发展,如人类特有的符号交流能力或发展投射武器的能力(参考文献19-23)。然而,现在已经证明复杂武器在非洲有相当长的历史(参考文献24、25),而且在海洋同位素5期(MIS 5,13-7.1万年前)期间,非洲(参考文献26-30)和西南亚(参考文献29、30)都已经出现了反复的符号行为标志。到约6万年前,东北非和邻近西南亚地区的中石器时代/中旧石器时代考古记录变得相当普遍(参考文献31、32),没有特别显著的、具有地区特征的共同特点。更广泛地说,非洲考古记录显示出复杂的、非线性的文化变迁轨迹(参考文献33、34)。这些变迁通常包括长期的停滞期,偶尔被不同非洲地区相对短暂的创新浪潮所打断(参考文献35-41),这发生在欧亚大陆创始人群出现前数千年(参考文献10、42)。考古记录中的大规模累积性变化也发生在6-5万年前之后(参考文献10),这表明现代人类在欧亚大陆的创始人群的成功并不依赖于某个特定的广泛技术创新或突然增加的认知能力(参考文献43)。
最近的研究集中在一系列相互关联的人口统计学解释上。特别是,非洲中石器时代考古遗址在12.5-5万年前的地理扩张被认为反映了人口可能增长,以及与主要扩张浪潮类似的祖先在非洲内部的扩张(参考文献1、44、45、46)。这种人口扩张据认为还包括更大的人口密度(参考文献47)和通过远距离社交网络增加的连通性(参考文献48),所有这些都可能作为"推动"因素。基于与古人类物种相关的生态位变化的定量研究(参考文献49),以及特定中石器时代石器工业(参考文献22),遗址的空间分布模式也被用来说明人类对"通才-专家"生态位的独特适应(参考文献50)。然而,这些假设都没有在大陆尺度上进行定量评估,很少有从合适理论模型的预测角度进行检验,这可能导致事后解释的泛滥(参考文献51、52)。因为人类在非洲内部和走出非洲的扩张可以与理想自由分布模型下的生态位宽度扩张联系起来(参考文献53),我们从人类生态位扩张的角度探讨了成功走出非洲的问题。随着人口密度增加,人类在更具挑战性的环境中定居变得有利可图。在这种条件下,当达到成功走出非洲所需的临界人口规模,且人类的生态适应能力增强时,扩张就可能发生(参考文献54、55)。
在这里,我们专门检验更新世智人的生态位在走出非洲之前是在非洲内部扩大还是收缩。我们将生态位定义为决定物种能够生存和繁衍的生物气候因素的集合(参考文献56)。为了回答这个问题,我们在前人研究的基础上(参考文献22、57、58、59、60),使用物种分布模型(SDM)方法(参考文献61)来测量更新世人类在非洲内部生态位宽度的变化。我们的结果经过两组不同古环境模拟的检验(参考文献62、63、64),正式证明人类生态位扩张先于并伴随着后来成功的走出非洲。
检验生态位变化
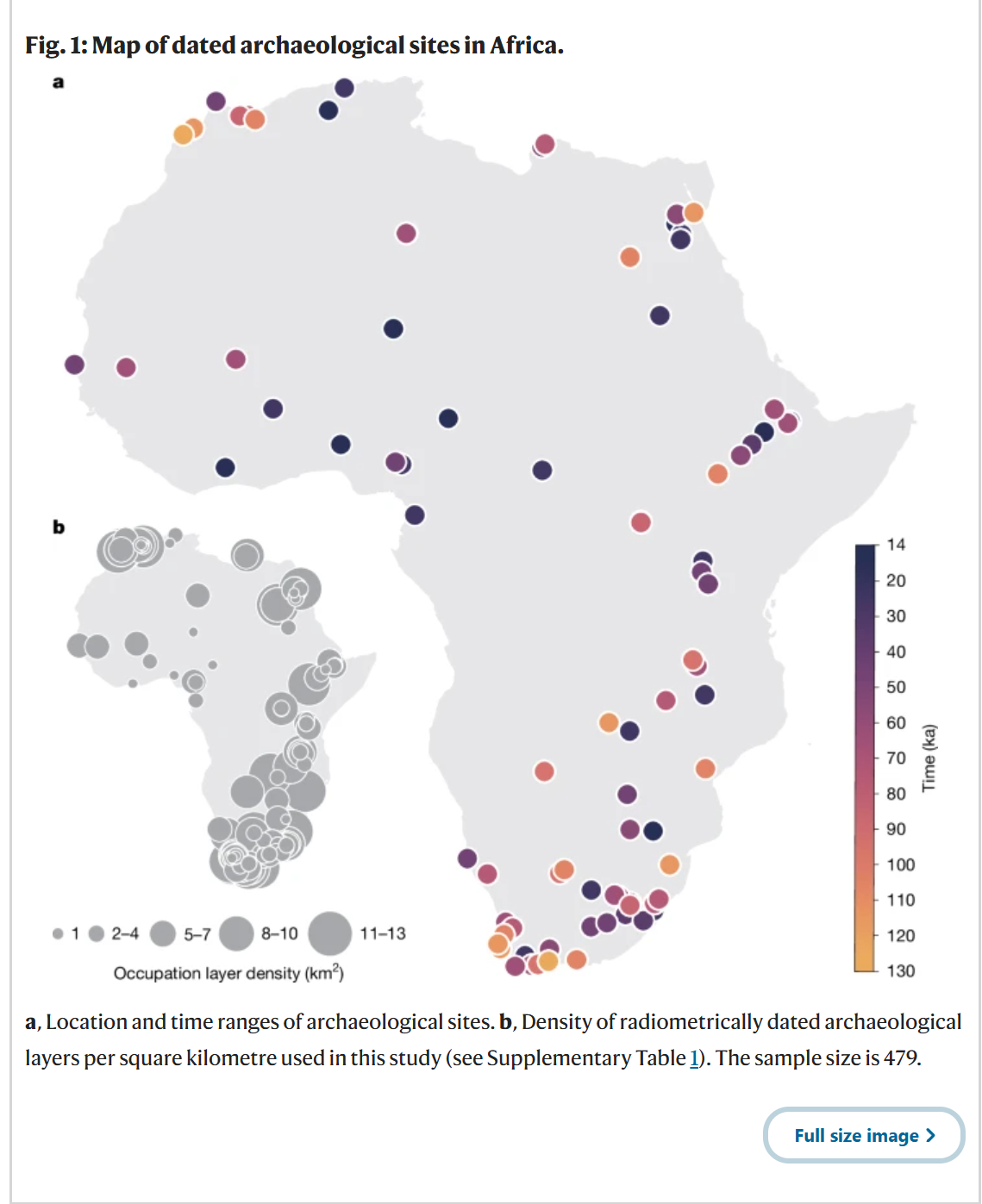
为了重建非洲大陆人类生态位随时间的变化,我们采用了泛非洲方法,建立了一个全面且精心策划的数据库,包含了年代在120-14千年前的更新世考古遗址的放射性测年的地层层位,并排除了年代不确定性大于20,000年的遗址(关于遗址选择的详细信息见方法,所包含的遗址见补充表1)。对于每个保留的地层层位(图1),它代表了一个确认的人类活动证据,我们提取了与其位置和年代相关的古气候变量和生物群系62(基于BIOME4模型的植被模拟),总共有479个放射性测年的记录。我们重点关注了五个生物气候和植被变量,这些变量似乎对于根据我们的数据重建栖息地适宜性最具参考价值(见方法):(1)叶面积指数(LAI);(2)年温度范围(BIO 7);(3)最湿季平均温度(BIO 8);(4)最暖季平均温度(BIO 10);以及(5)最湿季降水量(BIO 16)。由于每个特定生物群系的数据有限,BIOME4的植被模拟结果被合并为三个主要类别:森林、草原和沙漠(见方法和补充表4)。我们使用了两组古气候模拟:(1)哈德利中心耦合模式3(HadCM3(参考文献62)),其结果在正文中呈现,以及(2)瞬变社区地球系统模式(PCESM63,64),其结果在扩展数据图8a中显示。尽管这两组模拟有高度的相关性,但温度和降水在时空上的波动幅度有所不同(扩展数据图8a);尽管存在这些差异,所有关键结果在使用不同古气候模型时在质量上都是稳健的。
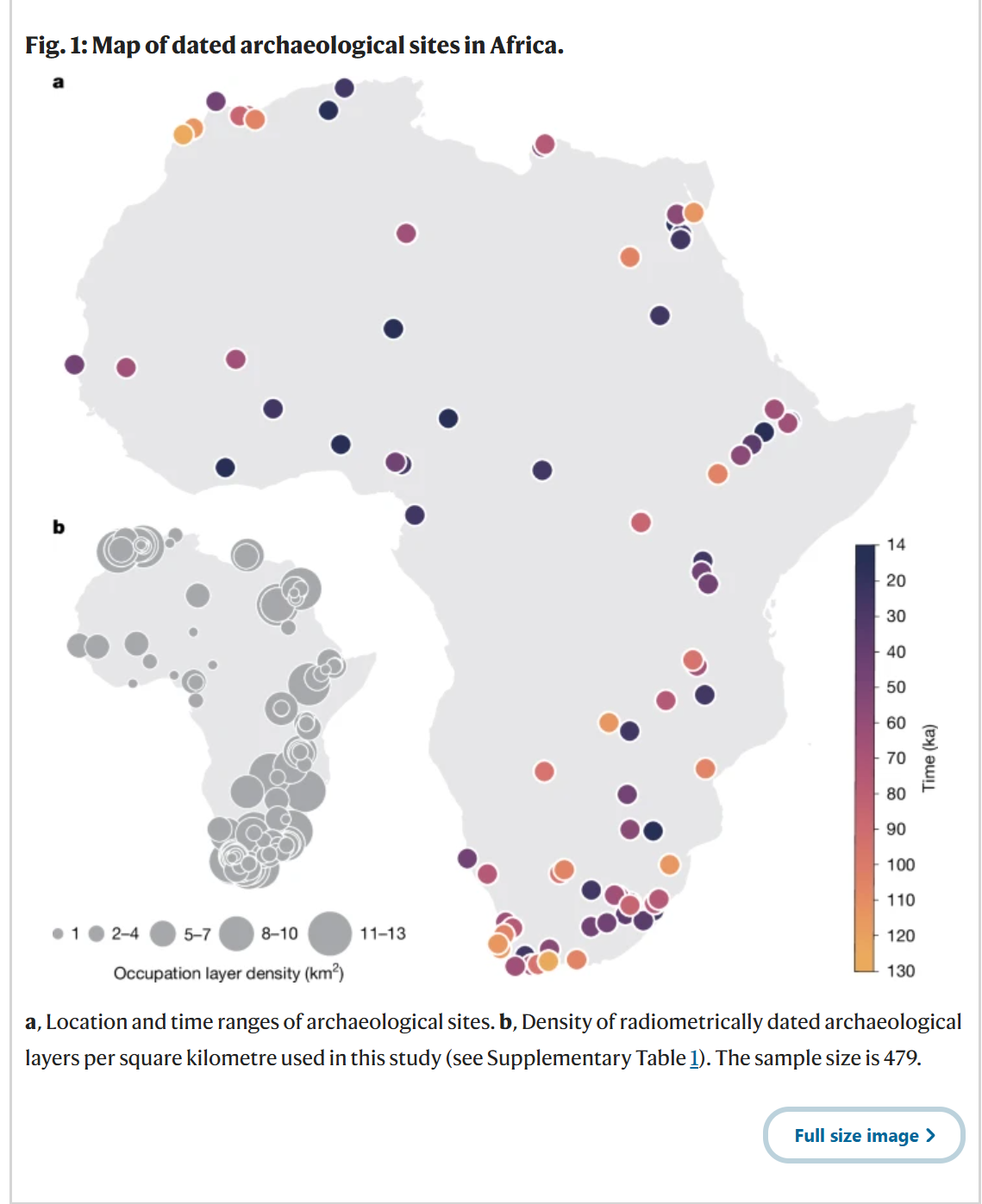
a, 考古遗址的位置和年代范围。b, 本研究中每平方公里放射性测年考古地层的密度(见补充表1)。样本量为479。
为了随时间重建人类生态位,我们使用了一种适配了五个生物气候变量的物种分布模型(SDM)方法。生物群落类型并未作为SDM的输入,而仅用于表征模型预测为适宜居住的区域的环境特征。传统上,通过将SDM拟合到不同时期并比较推断出的生态位来检验生态位变化。然而,更新世人类的数据非常稀少,无法创建具有足够样本的多个时间切片。相反,我们使用广义加性模型(GAMs)将整个时间序列作为单一数据集进行拟合。通过这种方法,可以通过比较两种GAM来正式检验生态位随时间的变化:一种GAM中每个环境变量对预测发生的影响在时间上保持恒定(固定生态位GAM),另一种GAM中该影响允许变化(变化生态位GAM;形式上,通过拟合每个预测变量与时间的交互作用;见方法部分)。
我们数据集中有年代的考古层数量随时间变化,越接近现代时期数量越多(Extended Data Fig. 1a)。这种模式可能是由于较近时期的遗迹保存机会更大,因此构成了一种可能扭曲结果的采样偏差。为了避免这种偏差,我们随机对观察进行二次采样,以生成随时间均匀分布的出现次数(Extended Data Fig. 1b)。我们重复此过程100次,生成几个"标准工作数据集",以探索这种二次采样的随机效应(Fig. 2)。我们按以下方式考虑了每个放射性测年相关的年代不确定性:在每个数据集中,我们将每次出现与从其整个可用年代范围中采样的日期关联起来(遵循正态分布,以平均值为中心)。最后,对于每个标准工作数据集,我们量化了气候变量相对于出现的背景分布,对每次出现采样200个按时间匹配的随机位置。接下来,我们将固定生态位GAM和变化生态位GAM拟合到每组标准工作出现次数及其相关的伪缺失点,从而得到我们分析的100次重复。使用赤池信息准则(AIC)进行的正式模型比较在100次重复中有91次强烈支持变化生态位GAM(见Methods)。为了理解模型检测到的生态位变化,我们将重复组合成一个集成66;具体来说,我们将AIC支持变化生态位GAM且Boyce连续指数(BCI;一种仅有出现数据的拟合优度度量)67大于0.7的重复组合在一起,并研究了随时间的变化。我们还从AIC < 2(因此不显著支持变化生态位模型)的重复中使用固定生态位GAM创建了一个集成,这为我们提供了一个在没有任何生态位变化情况下仅由气候波动引起的变化的零模型。在使用PCESM古气候模拟63进行相同分析时也可以观察到定性相似的模式。
图2: 智人按海洋同位素期(MIS)重采样出现次数。
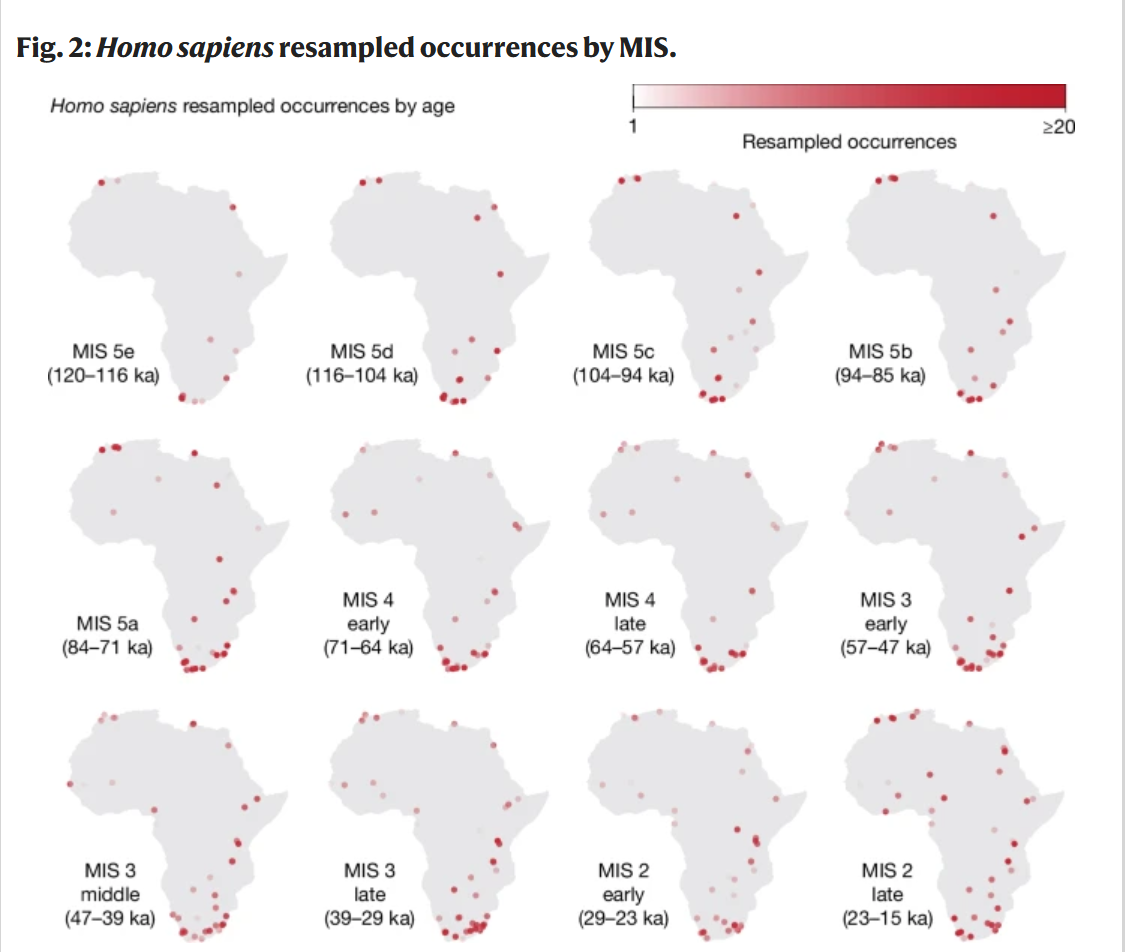
为了处理考古地层年代不确定性,我们从适当的年代范围内重新取样:点的颜色深浅表示每个海洋同位素期(MIS)中分配给特定位置的重采样次数(较深的点来自年代不确定性较低的地层,这些地层的大多数年代都落在同一个MIS内;较浅的点代表年代不确定性较高的地层,其重采样分布在多个MIS中)。有关重采样方法和MIS 4、MIS 3和MIS 2内部年代细分的详细信息,请参见方法部分。
栖息地范围在7万年前扩张
改变生态位的广义加性模型(GAM)集合显示,更新世人类在非洲的适居地理范围随时间显著增加(图3)。最显著的增加发生在大约7万年前MIS 4初期的西非、中非和北非地区。这次7万年前的大规模地理范围扩张之后,在MIS 2期间(约2.9万年前)又出现了一次覆盖非洲大部分地区的范围扩张(图3)。这些变化是生态位扩张的结果,因为固定生态位GAM预测整个大陆的人类占据范围相对稳定且广泛(扩展数据图2)。使用PCESM气候模拟进行相同分析也获得了定性上相似的结果(扩展数据图8b和10)。
图3:可变生态位模型中栖息地适宜性随时间的投影。
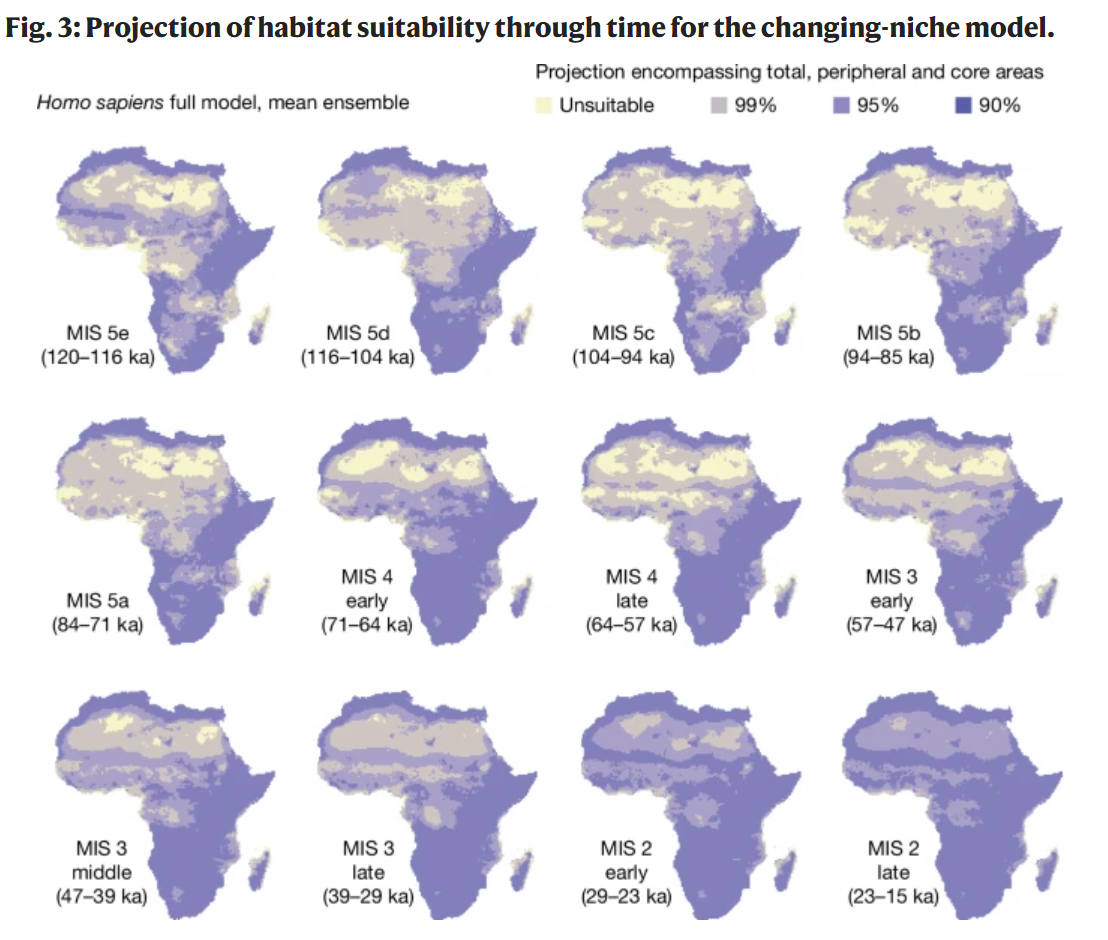
平均组合;详见方法。黄色代表不适宜区域,紫色深浅代表适宜性的高低。这些是作为总体区域、边缘区域和核心区域计算得出,分别包含了99%、95%和90%的考古发现点。每个MIS时期的地图是通过计算感兴趣时间段内每个单元格的栖息地适宜性平均值而生成的。MIS 4、MIS 3和MIS 2的年代学细分在方法部分有描述。所使用的气候数据来自Beyer等人(参考文献62),并加入了Heinrich事件。使用PCESM(参考文献63,64)得到的等效结果见扩展数据图8b。
生态位宽度的变化
为了更好地理解这些地理范围变化与由变化生态位GAMs集合所定义的实现生态位宽度之间的关系(图4a),我们研究了在变化生态位和固定生态位模型下每个生物群系类别的生态位宽度变化。我们对GAMs中使用的环境变量进行了主成分分析,将所有时间步骤汇集在一起,生成一个描述感兴趣时期气候变化的合成2D空间。对于每个时间步骤,我们然后测量在每个生物群系类别中由相应模型预测为适宜的位置上拟合的核心区域(图4b)。我们还探讨了变化生态位GAMs集合的相互作用图(即每个生物气候变量与时间相互作用的部分效应)(扩展数据图3),以了解生态位的确切变化,因为这些代表(理论上的)完整生态位,不受不同气候条件可用性的影响。图4b显示了非洲每个生物群系内实现的生态位大小,允许人类与气候关系发生变化(变化生态位模型),而扩展数据图4则将这种模式与假设这种关系保持恒定(固定生态位模型)的情况进行了比较(对于两个使用的古气候模拟)。因此,后者表示如何仅随气候波动而变化的人类分布。
图4:智人适宜栖息地和气候生态位面积随时间的变化。
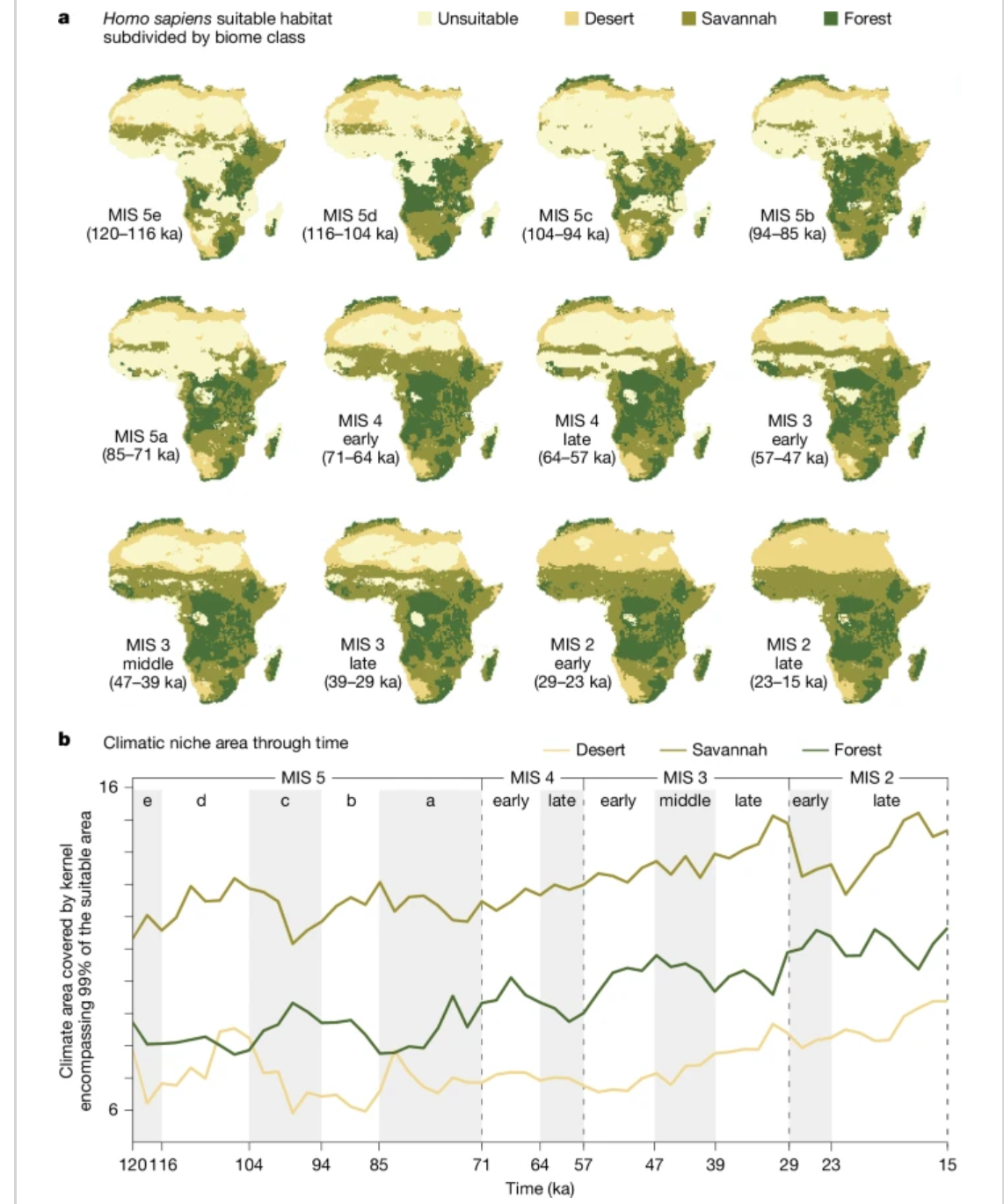
a,智人适宜栖息地(包括核心区和边缘区)按生物群系类别(沙漠、热带草原和森林)划分。b,气候生态位面积随时间变化。生态位面积是通过在气候变量的前两个主成分绘图上,估算包含每个生物群系类别适宜单元格(包括核心区和边缘区)的二维99%核密度来计算的。使用PCESM63,64得到的等效结果如扩展数据图4a所示。
如图4b所示,在MIS 5a之前,草原和森林的潜在使用量在相对稳定的水平上波动。在MIS 5a和MIS 4之间,这些栖息地的使用开始稳步增加。这种增加并非由于这些栖息地可用性的增加,因为在固定生态位GAM中并未观察到这种情况(见扩展数据图4a,与图4b相对应)。相反,最湿季节温度较高(BIO 8)且降水量较大(BIO 16;见扩展数据图3)的区域使用量增加,从而使生态位逐步扩展到森林。
沙漠的使用则表现出更大的变化,部分原因是可用沙漠的干旱程度存在波动。在较湿润时期,我们发现适合人类居住的沙漠生态位空间增加,因为它们的气候特征不那么极端。我们在固定生态位GAM的重建实现生态位面积中观察到这种效应,其中随时间的变化仅与影响可用气候条件的气候变化有关(见扩展数据图4a)。变化生态位和固定生态位GAM之间的关键区别在于,在前者中,在MIS 5a早期由于气候较为适宜而达到峰值后,沙漠的使用量保持在高于先前不利时期的水平。这种向沙漠的稳定扩张一直持续到MIS 3,那时我们看到沙漠栖息地的使用又出现了渐进式增长。这种情况在固定生态位GAM中并未观察到。
从机制上讲,沙漠使用量的增加是由于年温差较大的区域(扩展数据图3中的BIO 7)使用量增加,以及在LAI较低(即植被稀疏;扩展数据图3中的LAI)的区域生存能力逐步提高。尽管略有减弱,但在使用基于PCESM64的生物群系分类重复分析时也观察到了相同的模式(扩展数据图4b和9)。
讨论
我们的研究结果表明,人类生态位从约7万年前开始逐步扩张以包含更多栖息地类型,这种扩张在约5万年前达到顶峰,与成功走出非洲的时间相吻合。第二次扩张始于大约2.9万年前;到末次冰期第2阶段(MIS 2)时,人类已经占据了非洲的所有地区和生态系统,晚更新世的社会开始展现一系列新的行为,包括半定居生活、持续的大尺度社交网络的证据,以及领地意识和人际暴力的增加。我们的发现在使用两套不同且独立开发的古环境模拟时都保持一致。
从7万年前开始在非洲发生的人类生态位扩张,是由人类对森林和沙漠生物群落的偏好逐渐增加所驱动的,这使他们能够扩张到以前很少有人居住的地区:(1)西非森林,(2)中非森林,最终扩张到(3)干旱的撒哈拉地区和半干旱的萨赫勒地区。这种适应新栖息地的能力增强,从赤道森林到干旱沙漠的极端环境都能适应,使这些人类群体具备了生态灵活性,能够应对在走出非洲过程中遇到的各种新的环境条件,使他们在早期走出非洲失败的地方取得了成功。
因此,从7万年前开始在非洲发生的人类生态位扩张,为人类群体在约5万年前成功在全球扩张提供了解释,这与理想自由分布理论对人口扩张后果的预测也是一致的。
这些结果也为我们了解约7万至5万年前考古记录的变化提供了见解。在这段时期,不同地区呈现出独特的文化发展轨迹(参见补充信息)。特别是在这个时期及稍早时期的非洲,已有记录表明存在通过大范围焚烧进行的生态系统改造、水资源储存技术以及扩大的食物范围。这些发现可能反映了人类栖息地向新环境扩张的行为前提和表现。
需要强调的是,我们的泛非洲研究结果并不意味着整个大陆存在一个独特的、统一的进程。实际上,这些变化更可能与几个特定的文化复合体有关。我们需要开展更多研究来解开导致这些变化的原因。
我们强调,尽管我们记录到了生态位扩张,但这并不一定反映总体人口规模的增长,而且可能无法区分人口规模增长与生态位空间缩减条件下密度增加的影响。例如,由于承载力低,干旱地区的新人类生态位不太可能维持大规模或稳定的人口数量。相反,人类生态位的扩张表明人类能够在多个栖息地和地点之间迁移,这可能随着时间推移增加了不同群体之间的接触频率。如果情况确实如此,这或许可以解释为什么定义现代人类的一系列特征在10万到5万年前之间在个体中得以固定。这也为一个假说提供了独立支持,即在大约6万年前向欧亚大陆扩张的人群的祖先,可能在非洲内部经历过类似的扩张。
大约70-50千年前的时期在气候上是复杂的。虽然由于大西洋经向翻转环流减弱导致的海因里希事件6带来了整体更寒冷和干燥的时期,但这种影响在非洲各地区的程度不同。值得注意的是,代用记录显示在干旱和湿润时期之间反复转换,有迹象表明在海因里希事件6之后可能出现了影响大陆大部分地区的更广泛的湿润期。我们运行的HadCM3模型(结合Beyer等人和Armstrong等人的研究)捕捉到了这种时间和空间上的异质性;在没有明确包含海因里希事件6的PCESM模型中,这种模式较为微弱。
海因里希事件6对人类进化的影响主要被理解为其在人类走出非洲过程中所起的作用。有人认为北非和东非的干旱化可能作为推动力促进了人类迁出非洲;但也应该注意到,由于降温效应及其不均匀的影响,这一时期可能提供了有利于人类离开大陆的通道。类似的论点也适用于约29千年前MIS 2的开始,当时不均匀的干旱化促使人类进一步迁移到更干旱的栖息地。根据我们的研究结果,这些时期的气候不稳定可能有利于人类更广泛生态位的演化,进一步增强了人类应对区分智人属的多样化栖息地的能力。
通过在非洲建模从12万年前到1.4万年前的人类生态位,我们表明成功扩张到欧亚大陆并在那里建立长期人口的过程,本质上是始于非洲内部的一个过程。从根本上说,我们记录到了一个无可争议的非洲进程的开端,这个进程始于7万年前,最终造就了今天人类前所未有的生态适应能力。因此,我们的研究结果为理解和研究我们物种的全球扩散提供了新的基础。
 陕公网安备61011302002223号
陕公网安备61011302002223号