中国明清时期一个六代单系父系家族的基因组分析
[https://www.sciencedirect.com/science/article/pii/S2589004225012295](https://www.sciencedirect.com/science/article/pii/S2589004225012295) 1北京大学生物医学前沿创新中心,北京 100871,中国 2北京大学化学与分子工程学院,北京分子科学国家实验室,北京 100871,中国 3北京大学-清华大学生命科学联合中心,北京 100871,中国 4北京大学前沿交叉学科研究院,北京 100871,中国 5北京大学考古文博学院,北京 100871,中国 6北京市文物研
https://www.sciencedirect.com/science/article/pii/S2589004225012295
1北京大学生物医学前沿创新中心,北京 100871,中国
2北京大学化学与分子工程学院,北京分子科学国家实验室,北京 100871,中国
3北京大学-清华大学生命科学联合中心,北京 100871,中国
4北京大学前沿交叉学科研究院,北京 100871,中国
5北京大学考古文博学院,北京 100871,中国
6北京市文物研究所,北京 100009,中国
7深圳湾实验室细胞分析研究所,广东 528107,中国
要点:
- 通过古DNA重建了中国明清时期的一个六代家谱
- 严格的父系葬制,呈梯形空间分布
- 通过亲缘关系分析揭示了女性外嫁和一夫多妻制
- 前魏沟个体与黄河流域相关人群之间存在基因亲缘关系
**摘要:**历史时期的家族墓地通常遵循结构化的葬制模式,但由于文献记录有限和考古解释存在模糊性,识别这些安排具有挑战性。考古基因学提供了一种精确确定生物学亲缘关系的方法,使得社会关系和葬制习俗的重建成为可能。在本研究中,我们分析了北京明清时期前魏沟墓地的34位个体的古DNA和考古背景数据,重建了一个六代父系家谱。基因组数据揭示了严格的父系葬制,夫妻合葬,以及反映代际时序的非随机西向东空间分布。每一代都占据着不同的位置,形成梯形葬制模式。本研究展示了基因数据如何能够阐明历史上的亲属关系组织,完善对明清时期葬制习俗的假设,并加深我们对晚期帝制中国家族结构的理解。
引言:
家族墓地通常被定义为同一家族成员的墓葬,位于一个大致划定的区域内,并按有序方式排列和分布。在中国历史各个时期,考古发现表明家族墓地的葬制安排自西周时期就已存在多种形式,并从秦汉到魏晋时期发展成为系统性的实践。揭示家族墓地的布局可以帮助重建古代社会的家族组织性质,深入了解当时的各种社会和文化习俗。
然而,除了少数皇室或高级官员的高等级墓地外,许多可能代表普通民众的非特定墓地几乎没有详细的墓志铭或文献记录。在这种情况下,仅凭考古证据无法确定出土的个体之间的遗传关系。这些个体是否确实是血缘关系的同一家族成员仍然存在不确定性。因此,具体的葬制风格只能通过将人类遗骸的身体位置和方向与同时期的文献记录进行比较来推测,而无法做出确定的判断。
随着古DNA技术的进步,通过全基因组数据获取直接证据并确定同一墓地中个体之间的精确遗传亲缘关系已成为可能,从而可以重建家族谱系结构。结合考古和人类学发现,这些评估信息可以推断特定的葬制风格,进而反映当时社会的文化习俗。然而,迄今为止,关于中国历史时期生物学亲缘关系的古DNA研究仍是空白,这极大地阻碍了考古学和历史学对家族葬制的进一步探索。
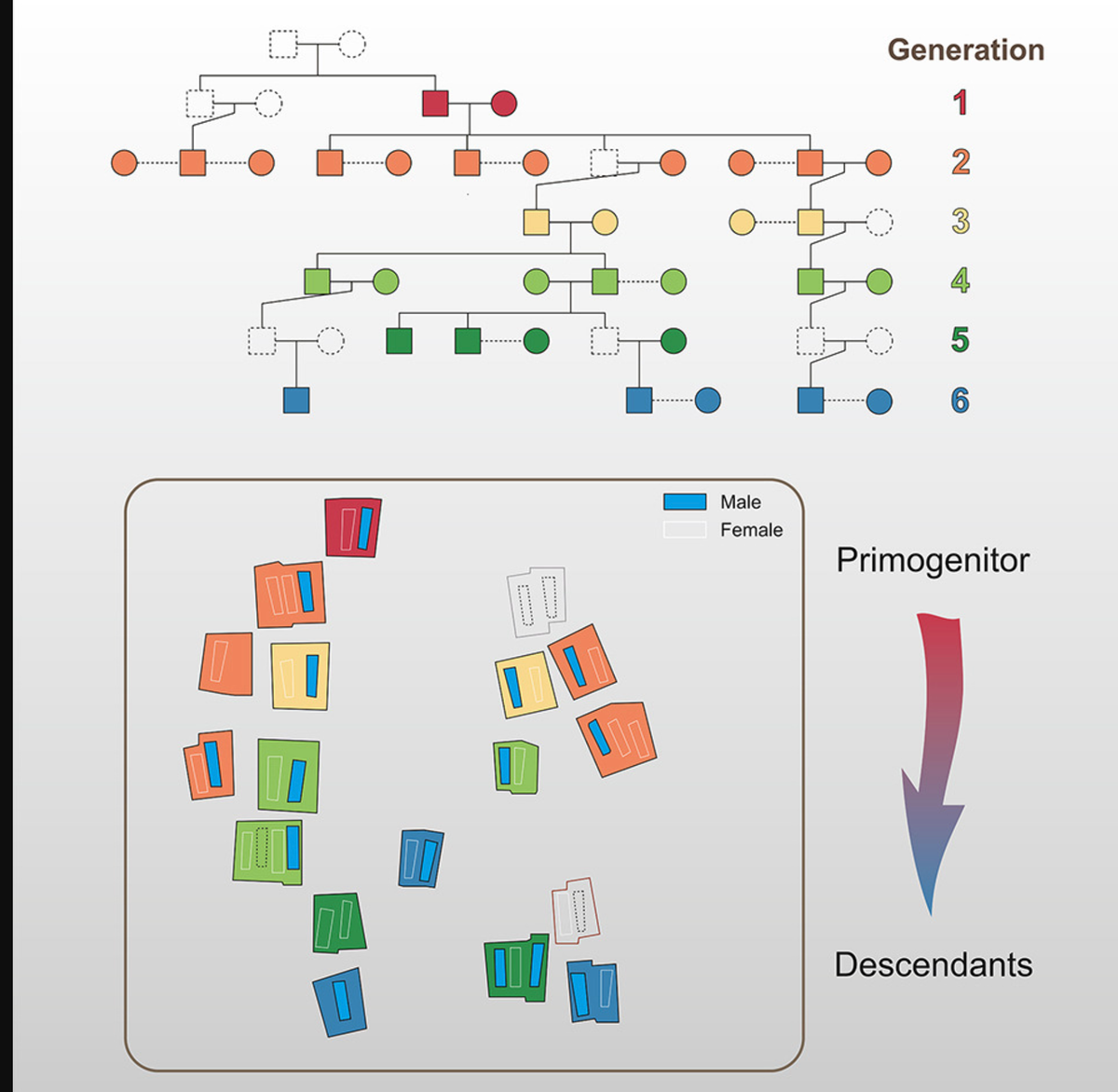
前魏沟遗址位于中国北京,考古学上归属于明朝(公元1368-1644年)和清朝(公元1644-1912年)时期。由于文物稀少且缺乏文献记录,仅凭考古证据几乎无法确定前魏沟墓地的具体葬制习俗。在本研究中,我们对该遗址出土的所有个体的人骨遗骸进行采样,获得了34个全基因组数据,以全面研究个体之间的遗传关系及其相关的葬制习俗。基因组证据,连同线粒体基因组和Y染色体数据,证实了该遗址存在一个至少延续六代的严格父系家族谱系。我们重建的家谱与墓葬的空间排列相吻合,表明这是一种经过刻意规划的葬礼,呈现出梯形排列的特定模式,这可能反映了当时普通家庭的文化特征。
结果
前魏沟遗址的考古和人类学见解
前魏沟遗址(北纬40°01′18.24",东经116°33′43.01")位于中国北京孙河乡东南部。该遗址由北京市考古研究所于2019年发掘,考古学上归属于明朝(公元1368-1644年)-清朝(公元1644-1912年)时期,包括19座竖穴墓(编号M1至M19),可分为四类:单棺(3座)、双棺(12座)、三棺(3座)和四棺(1座)(表1)。
其中,M1是一座位于发掘区西北侧的独立单棺墓(图S1);M2至M19集中分布在发掘区南侧,分别排列在墓地中轴线的南北两侧(图1A)。从M2至M19共出土34具人骨遗骸,头骨均朝向西方。个体根据墓号和在墓中的位置进行标记。例如,M3S、M3M和M3N分别代表M3号墓中位于北侧、中间和南侧的个体,其他墓葬也采用类似标记方式。
人类学评估显示,除M7、M8和M18号墓(可能由于后期扰动或盗墓导致缺失人骨)外,所有多棺墓均为男女合葬。大多数个体为成年人,只有M2S和M11M是约15岁的青少年女性。值得注意的是,男女埋葬的空间布局呈现一致性:在每座合葬墓中,男性埋葬在靠近墓地中轴线的一侧,而女性则位于外侧(图1A)。此外,通过考古发掘调查,通常可以确定合葬墓中个体的埋葬先后顺序,这为确定合葬墓中个体的死亡顺序提供了重要线索(图1B)。
除了人类遗骸,前魏沟墓葬出土的文物非常有限,仅包括少量陶罐、瓷罐和铜钱,这表明这是普通家庭的葬礼(图1A、1B)。大多数铜钱跨越了从明朝中期到清朝早期的连续时期,一些早期铜钱属于北宋时期(公元960-1127年)(图1A,数据S1)。可以通过这些铜钱的类型初步估计墓葬的年代顺序,因为墓葬的年代必然晚于最新铜钱的最初铸造时间。我们发现墓葬的时间顺序大致呈西向东排列,这一点还可以通过每座墓中人类遗骸头骨朝向西方的方位得到进一步证实。
古DNA鉴定和单亲遗传标记的多样性
前魏沟考古遗址的34位个体进行了全基因组测序,核基因组覆盖度从0.086×到2.384×不等,中位数覆盖度为0.877×。其中31位个体的核基因组覆盖度超过0.2×,30位个体的线粒体基因组覆盖度超过50×。对于剩余4个低覆盖度的个体(具体是M2N、M3M、M9M和M17S),使用人类线粒体DNA探针的靶向富集策略获得了完整的线粒体基因组数据。
古DNA的真实性通过多方面标准进行评估:显著的损伤模式和极低的现代人DNA污染率(表1,数据S2)。通过评估X染色体与Y染色体覆盖度的比率确定了34位个体的遗传性别,其中15位被确定为男性,19位为女性。
所有34位个体都获得了完整的线粒体DNA序列,覆盖度从33倍到2419倍不等,这些序列被归类为22种单倍型,通常与现代东亚单倍群相关。相比之下,所有15位男性个体都被归类为Y染色体单倍群D1a1b1a谱系(表1,图S2)。这种遗传特征表现出Y染色体多样性低而线粒体DNA多样性高的特点,表明前魏沟社区存在父系世系传承。
家谱重建及其与空间墓葬布局的关系
为了研究婚配模式,我们首先对34位个体的纯合片段(ROHs)进行了分析。我们的研究发现,只有六位个体表现出可检测到的ROH片段,其总推断ROH长度小于10厘摩(cM),且没有任何个体的单个ROH片段超过4 cM(图S3,数据S3A)。将这一观察结果与近亲繁殖和小群体内部通婚情况下预期的ROH长度理论分布进行比较,我们得出结论:前魏沟个体中没有发现这种做法的证据。
我们采用亲缘关系推断(KIN)方法来识别前魏沟个体之间的遗传关系,以阐明其亲缘结构。我们的分析发现了24对一级亲属关系,包括19对亲子关系和5对兄弟姐妹关系,以及28对二级亲属关系和21对三级亲属关系(图2A,数据S3B)。这些密切亲缘关系中的大多数(73对中的72对,占98.6%)要么是男性-男性(44对,占60.2%)要么是男性-女性(28对,占38.4%),而女性-女性关系(1对,占1.4%)明显较少。这一模式得到了成对错配率分析(图S4A,数据S3C)和性别特异性外群f3统计分析(Mubti;个体A,个体B)(图S4B)结果的证实。
基于推断数据对个体间共享的同源片段(IBD)分布的进一步分析也显示了一致的结果(图2A,数据S3D),所有这些都表明女性之间的遗传亲缘度最低,且显著低于男性之间的亲缘度。这些发现,加上Y染色体谱系的多样性低于线粒体DNA多样性的事实,可能暗示着该家族的女性来源更加多样化。
我们最初基于亲缘关系和单亲遗传标记以及死亡年龄和遗传性别的结果,构建了跨越两到三代的小型家谱。然后,我们逐步扩展这些家谱,形成了一个跨越六代的庞大家谱(图S5)。为了确定这个家谱树中两对二级亲属关系个体的具体类型(M12N和M10S以及M6S和M17),我们应用CREST21软件的性别推断模块来确定中间人的遗传性别。后续分析提供了有力证据,表明这两个中间人都是男性(数据S3E),说明M12N是M10S的父系祖父而非母系祖父,M6S和M17的情况也是如此。基于这一结果,可以确定M12S和M10S以及M6N和M17之间的亲缘关系为父系祖母-孙子关系。
我们对另外两对具有二级亲属关系的个体(M15N和M3S以及M12N和M14S)的各种亲缘关系组的IBD片段分布模式进行了研究。结果显示,它们与其他叔侄关系的个体对聚集在一起(图2B)。结合考古证据,即M3号墓中出土铜钱的年代大致与M4和M19号墓相当但晚于M15号墓,以及M15N和M3S之间不同的线粒体DNA单倍型,我们推断M3S很可能是M15N的侄子,而不是M15N的长辈亲属。同样,由于M14S与M12N的祖父母合葬,排除了M12N是M14S长辈的可能性,因此推测M14S是M12N的姨母。
此外,通过检验代际关系,我们为所有六对有子女的男女配偶是合葬在同一墓中的夫妻提供了确凿的遗传学证据,例如M15N和M15S、M14N和M14M、M5S和M5N以及M11N和M11S等配对,这也证实了长期以来关于夫妻合葬习俗的考古学假设。因此,剩余10位没有直系后代的女性个体可以根据上述推断填入这个家谱结构中,最终重建了包含33个个体的完整家族树(图3)。
这个家谱展现了清晰且连续的父系世系,其中所有男性成员都共享一个共同祖先(M15N)。这个大家族起源于M15N和M15S,他们有四个儿子,大多数家族成员都来自他们的两个儿子M14N和M16S的配偶。从西向东的埋葬顺序按时间顺序从祖先到后代进行,每个合葬墓中的男性都埋葬在靠近墓地中轴线的一侧,这表明空间布局从一开始就经过了精心规划。我们发现每一代直系后代的墓葬都沿着中轴线南北两侧的外缘排列。这种特定的埋葬模式可能属于历史文献中记载的"梯形排列"类型,反映了当时平民家庭的文化特征(图3)。
具体而言,第一代M15占据墓地最西端的位置,他的四个直系后代(M14、M4、M16、M19)和一个侄子(M3)位于南北两侧的外缘。M8中缺少人类遗骸可能是由于近期挖掘坑造成的干扰,导致骨骼损失和损坏。考虑到其在北部外缘的位置,M8原本埋葬的个体很可能与M15有家族关系,可能是他的儿子。后续几代——包括第三代(M13、M5)、第四代(M6、M12、M11)和第五代(M18、M9)——都埋葬在墓地的内侧,遵循着从西到东的分布方案。然而,第六代的埋葬位置出现了轻微偏离,M17和M6的墓葬位于西端,而M2的墓葬则位于中轴线上。尽管M2墓占据中轴线位置,考古分析显示M2的埋葬习俗和陪葬品与其他墓葬并无显著区别。因此,我们推测M2的中心位置不太可能有特定目的,如强调其优越的家族地位或财富。相反,其空间布局的不规则性可能源于有限的埋葬空间,或后代在执行埋葬习俗时的随意性。
前魏沟个体的遗传祖源和外貌特征
为了阐明前魏沟人群的遗传特征,我们使用主成分分析(PCA)22来研究古基因组与现代欧亚人群遗传多样性之间的关联。结果显示前魏沟个体与东亚人群聚在一起(图S6)。值得注意的是,在现代东亚人群的PCA分析中,前魏沟个体与现代北方汉族人群的关系比与南方汉族人群更为密切。他们在中国北方(YR_LN和YR_LBIA)23和中国东北辽西地区(WLR_LN和WLR_BA)23的古代人群之间形成了一个独特的聚类,这些人群年代从新石器时代晚期到青铜铁器时代(图4A),表明北方中国人群对前魏沟人群有遗传影响。
为进一步研究前魏沟人群与东亚人群的遗传联系,我们采用外群f3统计19,格式为f3(前魏沟, X; Mbuti),以Mbuti作为外群(图S7,数据S4)。我们的发现表明,前魏沟人群与庙子沟_MN、WLR_LN、YR_LN和YR_LBIA等古代人群表现出最高的遗传漂移,说明与黄河相关人群有显著的祖先遗传成分共享。
此外,我们进行了f4统计分析,格式为f4(Mbuti, Y; X, 前魏沟),将81个古代和现代东亚人群同时作为Y和X(图S8,数据S5)。这种方法通过统计显著的Z值直接比较前魏沟人群与81个古代和现代东亚人群之间的遗传差异,显著结果较少表明前魏沟与人群X之间的遗传相似性更强。我们在Z值分布中发现了明显的区域聚类模式,来自黄河(YR)和西辽河(WLR)地区的人群(X)产生了更多非显著的Z值,突显了它们与前魏沟人群相比其他东亚人群具有更强的遗传亲缘关系(图4B)。
最有说服力的证据来自前魏沟与YR_唐朝人群24的比较,即f4(Mbuti, Y; YR_唐朝, 前魏沟) (-2.703 < Z值 < 3.008),98.8%(79/80)的东亚人群未超过显著性阈值|Z值| > 3。这表明几乎所有分析的东亚人群都与前魏沟和YR_唐朝人群共享相等的遗传漂移,进一步表明它们之间最密切的遗传亲缘关系。
为揭示前魏沟人群中远古混合的潜在痕迹,并通过与现存人群的比较阐明该地区过去四个世纪中人类关键表型特征的可能变化,我们进一步研究了前魏沟基因组中与关键表型相关的SNP。我们首先使用HIrisPlex-S25-27,该系统使用41个SNP位点的二倍体基因型来预测与色素沉着相关的外部可见人类特征,包括头发、皮肤和虹膜颜色。在具有足够测序覆盖度用于每个特征表型预测的前魏沟个体中,所有人都被预测具有棕色眼睛(16/16)、黑色或棕色头发(20/20)以及中等、深色或深黑色皮肤(24/24)(数据S6)。男性和女性个体之间的表型没有显著差异。
这些结果表明,前魏沟个体统一展现了当地现代人群典型的东亚外貌特征。此外,我们分析了具有足够测序覆盖度的前魏沟个体在5个东亚人特有的特征相关SNP28-34上的基因型(数据S7),预测结果与现代东亚人的等位基因频率35,36一致。绝大多数前魏沟个体可能具有更直、更粗的头发和铲形门齿(29/30)、干耳垢和无体臭(24/26)、乳糖不耐受(7/8)以及无酒精冲红(24/28),这些都是现存当地人群的典型表型。总的来说,上述结果显示该地区人群在过去几个世纪中具有遗传连续性。
讨论
严格的父系葬制及特定空间格局
前魏沟考古遗址展示了一个清晰的父系葬制系统,其特点是一个六代单系家族结构,所有男性之间都保持着直系血缘关系。与稳定的父系血缘形成鲜明对比的是,前魏沟墓地中的女性被认为来自多个外部来源。这些女性只与她们自己的男性后代有关联,而与同代的男性没有关系。此外,我们没有观察到姐妹、兄妹、父女或母女等遗传关系,表明前魏沟遗址中没有埋葬女性后代。结合上述有力证据,这个家族很可能实行女性外嫁制,这是明清时期最普遍的婚姻制度。
前魏沟家族墓地的布局相对有序。考古学家通常认为这属于昭穆制埋葬模式,这是明清时期华北地区家族墓葬中的常见做法。这种葬制起源于周朝,其特点是按世代顺序排列,从最早的祖先开始,通常每代人排成一行(或一列),每代人依次排列。但是,通过基因亲缘关系分析表明,前魏沟家族的埋葬安排并不符合昭穆制模式。相反,它更接近于"卧马"葬制模式,即每代人的墓葬呈对角线排列,整体形成梯形的V字形图案,族长的墓葬位于顶点,类似于飞行中的大雁队形。这种模式有效地解释了为什么前魏沟家族墓地中同代人的几座墓葬并不位于同一行或同一列。尽管没有墓志铭或其他信息来确定祖先墓的位置,但古DNA结果已经明确了其位置,确定了所有成员之间的亲缘关系,并确定了每一代的具体位置,从而证实了实际使用的埋葬模式。因此,在许多已发表的明清时期家族墓地相关资料中,所描述的昭穆制埋葬模式可能并不准确,需要借助DNA测序等方法进行进一步澄清。
婚姻形式和亲属关系的多样性
在前魏沟的19座墓葬中,有16座被确认为合葬墓。通过确认后代之间的遗传关系,这些合葬墓很可能是用于已婚夫妇,其中大多数为一夫一妻制,因为16座墓中有12座包含两具棺材。此外,还有3座三棺墓和1座四棺墓存在多位女性配偶。根据考古发掘和死亡年龄的调查,可以区分这些墓葬中不同个体的埋葬顺序,这可能表明他们的婚姻是一夫多妻制,还是在原配偶死后的再婚。
例如,在M11墓中,男性个体M11N被确认比女性M11M和M11S更早埋葬,这排除了sequential婚姻的可能性,表明两位女性是同时与M11N结婚的一夫多妻关系。相比之下,在M3墓中,M3M被确认为成年女性,比中年女性M3N和男性M3S更早埋葬。一种可能是M3S同时有两位配偶,另一种可能是M3S在M3M死后与M3N结婚。我们没有进一步的证据来推断或排除这两种可能性中的任何一种。
在三棺墓M9中,我们出乎意料地发现了两名男性和一名女性,这与其他包含多位女性配偶的墓葬不同。M9N和M9S被确认为中年男性兄弟,这可能表明存在嫂婚(即一个兄弟娶另一个兄弟的遗孀)的可能性。然而,根据考古证据,M9M被确认为年轻成年女性,且比两兄弟更早下葬,这使得这种婚姻形式的可能性不大。此外,这种婚姻安排在历史上的中国非常罕见,通常只在一些少数民族中实行。明清时期占主导地位的婚姻制度是一夫一妻制,多配偶制主要表现为纳妾的形式。因此,我们缺乏实质性证据支持前魏沟家族允许嫂婚的说法。此外,从这3具棺材的空间布局来看,很可能只有M9S和M9M是安排在合葬墓中。作为M9S的兄弟,M9N被埋葬在其附近是合理的,但埋葬位置有所偏离,导致了不正常的接近和混淆。
前魏沟的墓葬还呈现出几种独特情况。首先,发现了几座单棺墓,包括M16和M17墓。根据家谱,M16墓北部缺失的个体是M15N的儿子,并与M16S育有后代。鉴于记录显示M16墓曾被现代水管系统破坏,我们推测这位配偶曾被埋葬在这座墓中,但其遗骸可能已经丢失或被破坏。M17墓可能代表了另一种情况,因为它包含一位中年男性的埋葬,这表明他可能在结婚前就去世了。
我们还在家谱中发现了一个不寻常的二级亲属关系,即M14S(第二代M14N的妻子)与M12N(第四代M14N的孙子)之间的姑侄关系。具体来说,M14S嫁给了M14N,而她的姐妹则嫁给了M14N的儿子M13N,并生下了儿子M12N。这种情况可以简化为一对姐妹分别嫁给父子二人。这种婚姻不被视为乱伦,而是跨代婚姻,因为他们之间没有直系血亲关系,也不存在遗传或基因因素。因此,这不属于近亲婚姻。我们为明末清初时期存在跨代婚姻提供了基因证据,反映了那个时代多样化的婚姻状况。
 陕公网安备61011302002223号
陕公网安备61011302002223号